the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 16 Dec 2019
| 16 Dec 2019
Latest Oligocene to earliest Pliocene deep-sea benthic foraminifera from Ocean Drilling Program (ODP) Sites 752, 1168 and 1139, southern Indian Ocean
Dana Ridha
Kirsty M. Edgar
Deep-sea benthic foraminifera provide important markers of environmental conditions in the deep-ocean basins where their assemblage composition and test chemistry are influenced by ambient physical and chemical conditions in bottom-water masses. However, all foraminiferal studies must be underpinned by robust taxonomic approaches. Although many parts of the world's oceans have been examined, over a range of geological timescales, the Neogene benthic foraminifera from the southern Indian Ocean have only been recorded from a few isolated sites. In this study, we have examined 97 samples from Neogene sediments recovered from three ODP sites in the southern Indian Ocean (Sites 752, Broken Ridge; 1139, Kerguelan Plateau; 1168, west Tasmania). These data cover a range of palaeolatitudes and water depths during the Miocene. More than 200 species of benthic foraminifera were recorded at each site and, despite their geographic and bathymetric separation, the most abundant taxa were similar at all three sites. Many of these species range from late Oligocene to early Pliocene demonstrating relatively little faunal turnover of the most abundant taxa during the key palaeoclimatic shifts of the Miocene. We illustrate and document the occurrence of the 52 most abundant species (i.e. those with >1 % abundance) encountered across the three study sites.





The Neogene can be informally subdivided into an early warm interval and late cool interval (Flower and Kennett, 1993). Superimposed on this long-term cooling is the Mid-Miocene climatic optimum (MMCO) between ∼15 and 17 Ma, marking the warmest interval of the Neogene (Zachos et al., 2001). Cooling following the MMCO culminated at ∼ 13–14 Ma in major growth of the East Antarctic Ice Sheet (EAIS). Further cooling coincided with increasing intensification of Antarctic deep-water formation and growth of the AIS and Greenland ice sheet (Flower and Kennett, 1993, 1994, 1995).
Profound palaeoclimatic and palaeoceanographic changes in the Neogene drove progressive and significant turnover of deep-sea benthic foraminiferal communities from the so-called transitional fauna characterising the late Oligocene–early Miocene to the late Neogene fauna of the late Miocene and finally the establishment of recognisably modern assemblages in the mid-Pleistocene (Jones, 1994; Kaiho, 1994). The first major transitional interval occurred in the early-to-middle Miocene, between ∼13–17 Ma (Kaiho, 1994), spanning the onset of the MMCO, subsequent East Antarctic Ice Sheet growth, and associated changes in water mass character and upwelling intensity (Woodruff, 1985). Further changes in assemblages that occur in the late Miocene, ∼8–10 Ma, relate to global cooling and expansion of the Antarctic ice sheet (Gupta et al., 2004). Finally, the last global extinction of benthic foraminifera (∼20 % of deep-sea genera) occurred in the late Pliocene–mid-Pleistocene (Hayward et al., 2012) and is variously attributed to increasing ocean productivity and intensification of low-oxygen zones and changes in the food type and supply, due to the expansion of the Antarctic and later Northern Hemisphere ice sheets and global cooling (Hermoyian and Owen, 2001; Gupta et al., 2004; Smart et al., 2007).
Superimposed on this long-term global pattern of benthic foraminifer assemblage change are distinct basinal and latitudinal differences in the timing of certain biotic events. For instance, the global decline in deep-sea benthic foraminifer species abundance and richness occurs much earlier (late Oligocene–Miocene) in the Southern Ocean than in the Indian Ocean (late Miocene; Hayward et al., 2012). However, the response of deep-sea benthic fauna in the Indian Ocean to these global changes remains relatively poorly constrained.
Previous records of Miocene benthic foraminifera from the central and southern Indian Ocean are largely single-site reports with a focus on palaeoceanographical interpretations, though some provide useful illustration for comparison with the present study. Boersma (1990) reported benthic foraminifera from 12 ODP sites from leg 115 across the central Indian Ocean, where quantitative studies of the benthic foraminifera indicated faunal separation at intermediate, deep and abyssal water depths from the late Oligocene onwards, and hence the development of a more strongly vertically stratified ocean. Nomura (1991) systematically described the most abundant Oligocene-to-Pleistocene benthic foraminifera species and assemblage changes at ODP Sites 754 (Broken Ridge) and 756 (Ninetyeast Ridge) in the eastern Indian Ocean, linking them with palaeoceanographic changes.
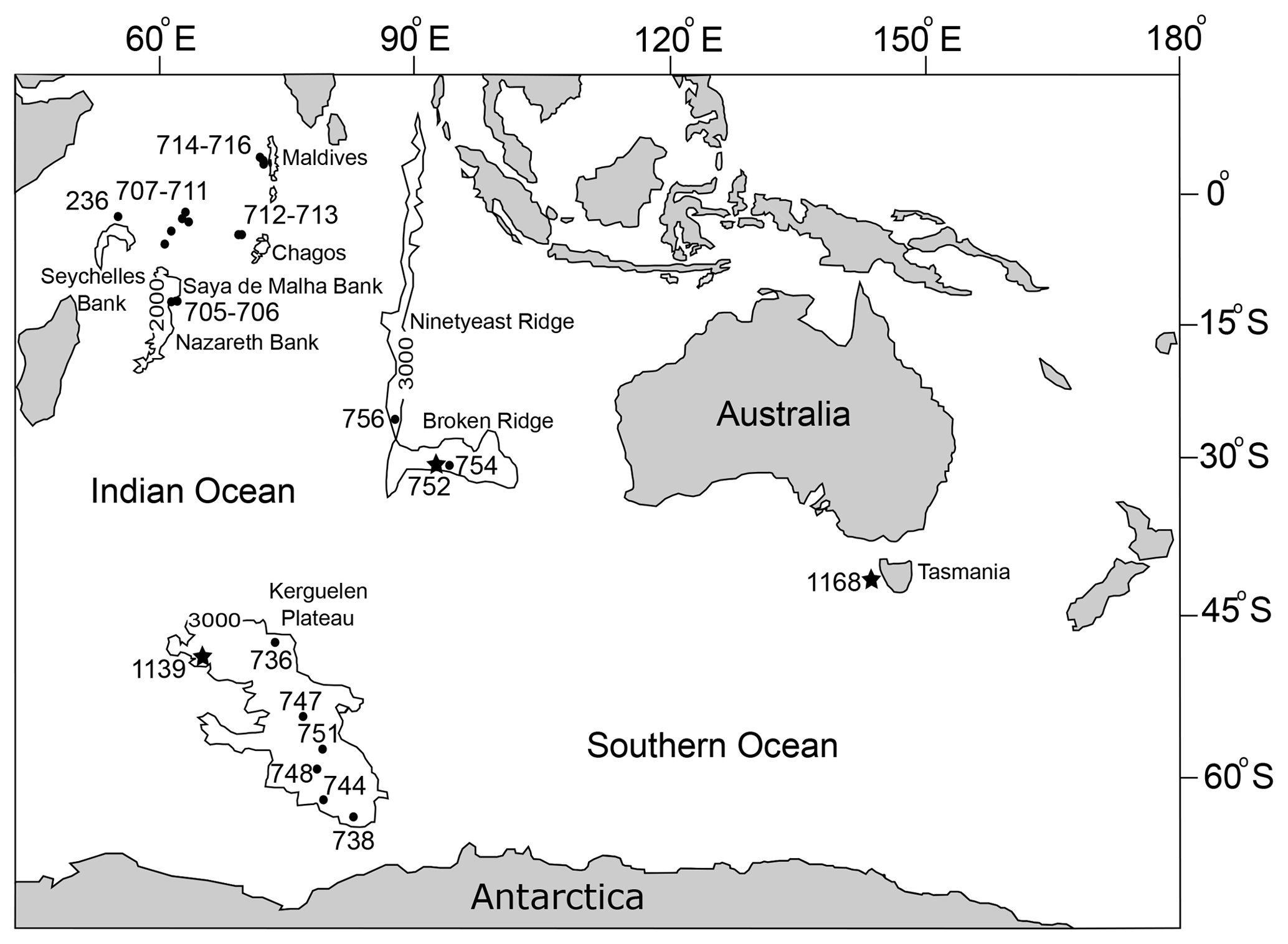
Figure 1Location map of the study area. Studied ODP sites are marked as stars (752, 1168 and 1139), additional sites referred to in the text are located by dots (after Weatherall et al., 2015). Sites 752, 754 and 756 (leg 121), Site 1168 (leg 189), Site 1139 (leg 183), Sites 705–716 (leg 115) and Deep Sea Drilling Project (DSDP) Site 236 (leg 24). The isobath adjacent to the Kerguelen Plateau, Broken Ridge and Ninetyeast Ridge is 3000 m, while the isobath near the Maldives, Chagos, Saya de Malha, Nazareth and Seychelles banks is 2000 m.
There have been two studies focussing on the Kerguelen Plateau. Schroder-Adams (1991) investigated middle-Eocene-to-Holocene benthic foraminifera from Sites 736, 738 and 744 on the Kerguelen Plateau (Fig. 1), describing and illustrating many species. Significant faunal changes (and species replacements) were noted in the middle Eocene, late-Eocene–early-Oligocene, middle Miocene and late Miocene. Mackensen (1992) quantitatively studied benthic foraminiferal assemblages from the lower Miocene through to upper Pleistocene interval from ODP Site 747 (Central Kerguelen Plateau) and Sites 748 and 751 (Southern Kerguelen Plateau).
Gupta and Satapathy (2000) studied abyssal benthic foraminifera from 31 samples of the latest Miocene to Pleistocene from Deep Sea Drilling Project (DSDP) Site 236, north of the Mascarene Plateau. This study investigated the link between benthic foraminifera distribution, productivity and deep-ocean thermohaline circulation in the west-central Indian Ocean during that period.
The main aim of this work is to provide a taxonomic and stratigraphic reference for the most common Miocene benthic foraminifera encountered in the southern Indian Ocean, supported by scanning electron microscopy (SEM) images. The study deals with foraminifera encountered in ODP Holes 752A, 1139A and 1168A, all in the southern part of the Indian Ocean (Fig. 1). Of these, the latter two have not previously been investigated for benthic foraminiferal assemblages; whilst Neogene benthic foraminifera assemblages have been used for palaeoceanographic reconstructions in Hole 752A (Singh et al., 2012), no systematic species description or reference images were provided. The present study expands our understanding of the spatial and temporal distribution of the most abundant benthic foraminifera from the deep-sea Miocene sediments of this region.
ODP Site 752 is situated on the Broken Ridge in the south-eastern Indian Ocean (30∘53.475′′ S, 93∘34.652′′ E; Fig. 1) at a current water depth of 1086 m (Shipboard Scientific Party, 1989). The palaeodepth reconstruction for the site is upper bathyal in the late Oligocene shallowing to mid-lower bathyal in the Miocene (Shipboard Scientific Party, 1989). The current study covered a 79.7 m interval of sediments from Hole 752A, which includes the latest Oligocene through to early Pliocene ages; the average core recovery is 70.6 %, composed mainly of foraminifer and nannofossil ooze. An age model for Hole 752A was developed using the calcareous nannofossil datums from the initial report (Shipboard Scientific Party, 1989), with revised ages from the Astronomically Tuned Neogene Time Scale (ATNT2012) in GTS2012 (Hilgen et al., 2012).
ODP Site 1139 is located west of Kerguelen Island (50∘11′20′′ S, 63∘55′40′′ E; Fig. 1) with a modern water depth of 1415 m consistent with a bathyal palaeodepth throughout the late Oligocene to earliest late Miocene (Shipboard Scientific Party, 2000). The late Oligocene to earliest late Miocene interval was sampled from Hole 1139A, comprising 167.3 m with average core recovery of 62 %. The sediments are foraminifera-bearing nannofossil ooze, nannofossil-bearing clay, and claystone and nannofossil-bearing ooze and chalk. An age model has been established using planktic foraminifera (Shipboard Scientific Party, 2000) and calcareous nannofossil datums (Persico et al., 2003) with ages revised using ATNT2012 in GTS2012 (Hilgen et al., 2012).
ODP Site 1168 is situated at a middle bathyal water depth (2463 m) on the western margin of Tasmania (42∘38′40′′ S, 144∘25′30′′ E; Fig. 1) (Shipboard Scientific Party, 2001). The sedimentary interval studied in Hole 1168A covers 422.8 m with an average recovery of 98 %. This interval covers the latest Oligocene through to early Pliocene, which is mainly composed of nannofossil ooze, silty clay and claystone. The age model for Hole 1168A uses planktic foraminifera, calcareous nannofossils and geomagnetic polarity datums calculated by Stickley et al. (2004) with revised ages from ATNT2012 in GTS2012 (Hilgen et al., 2012).
In total, 97 samples were selected from the study sites (30, 25 and 42 samples at ODP Sites 752, 1139 and 1168, respectively). Sediment samples were processed by first disaggregating them in water and then wet sieving at 63 µm. Samples were then dried overnight in an oven at 40 ∘C, the dry >63 µm sieved size fraction was then split until an aliquot of >300 benthic foraminifer specimens was achieved and subsequently picked clean of all benthic foraminifer. The 52 most abundant species encountered (of more than 500 species provisionally determined across all three sites) were selected for this work as they represent ∼79 % of the total benthic foraminifera specimens found across all sites. Scanning electron microscope images of representative (uncoated) specimens of each selected species were taken on a Phenom Desktop SEM at the University of Birmingham. The total size range of specimens observed within the study was also recorded by measuring the longest axis of the specimens using a reticle on a stereo microscope.
In the current study the generic and suprageneric classification of the foraminifera follows Loeblich and Tappan (1988, 1994). The classification of Loeblich and Tappan (1994) is broadly the same as that of Loeblich and Tappan (1988) with some modification of high-level categories as given in Loeblich and Tappan (1992). Specifically, the elevation of the foraminifera to a higher taxonomic rank by Lee (1990) to the class Foraminiferea was accepted by Loeblich and Tappan (1992) and Kaminski (2004). The key sources for species identification are Wright (1978), Hayward, (2002), Hayward et al. (2012) and Holbourn et al. (2013). Descriptions are based on existing literature with additional observations from this study.
Benthic foraminifer assemblages are relatively well-preserved with specimens largely complete and surficial details preserved. Assemblages are diverse with more than 200 species identified in total in each site (500 within the entire study) and are dominated by calcareous benthic foraminifera. This diversity is higher than previously documented in the Indian Ocean (e.g. Boersma, 1990; Nomura, 1991; Mackensen, 1992; Schroder-Adams, 1991). The 52 selected species described here are assigned to 33 different genera and are subdivided into four orders, 15 superfamilies, 20 families and 16 subfamilies. Four recorded species are left in open nomenclature, two each from Bolivina and Gyroidinoides. All taxa described are calcareous hyaline forms with the exception of one agglutinated species. Representatives of the Buliminida (31 species) constitute the majority of those recorded, especially in ODP Hole 752A, but they are also both abundant and diverse in Holes 1139A and 1168A. Species of the Rotaliida are the next most important components of the benthic foraminifer assemblages.
-
Class Foraminifera J. J. Lee, 1990
-
Order Buliminida Fursenko, 1958
-
Superfamily Bolivinoidea Glaessner, 1937
-
Family Bolivinidae Glaessner, 1937
-
Genus Bolivina d'Orbigny, 1839b
-
Bolivina dilatata Reuss 1850
-
Figure 3: 1a–b
-
1850 Bolivina dilatata Reuss: p. 381, pl. 48, fig. 15a–c
-
1949 Bolivina subspathulata Boomgaart, p. 112, pl. 12, fig. 4
-
2016 Bolivina dilatata Reuss; Lei and Li, p. 202, pl. 2, figs. a–h
Description
The test is biserial elongate, has perforate hyaline wall, the shape of the chambers is triangular to trapezoidal. The sutures are depressed. The aperture is a slit-like to arcuate opening with a thin tooth plate in the middle. The aperture location is terminal.
Dimensions
The species length ranges from 125 to 250 µm in the current study.
Stratigraphic range
This species is recorded in the Aquitanian of 1139 (ODP Site 1139) and late Oligocene (Chattian) to early Pliocene (Zanclean) at 1168. This species is not recorded at 752, consistent with earlier studies (Nomura, 1995).
Remarks
This is considered a cosmopolitan species (Lei and Li, 2016) but to the best of our knowledge this is the first time it has been recorded in the Indian Ocean.
-
Bolivina huneri Howe, 1939
-
Figure 3: 2a–b
-
1939 Bolivina huneri Howe: p. 66, pl. 9, figs. 3, 4
-
1983 Bolivina huneri Howe; Tjalsma and Lohmann, p. 23, pl. 11, fig. 5
-
1987 Bolivina huneri Howe; Miller and Katz, p. 124, pl. 1, figs. 7a–b
-
2013 Bolivina huneri Howe; Holbourn et al., p. 68, figs. 1–3
Description
Very densely ornamented biserial species with tapered test. The test surface has a reticulate ornamentation largely obscuring sutures with the exception of the final chamber. The inflated chambers are elongated. The aperture is situated at the base of the last chamber and is arcuate with wide aperture view and tooth plate.
Dimensions
The species length ranges from 175 to 250 µm in the current study.
Stratigraphic range
This species is recorded from the Chattian through to early Burdigalian of 752. At 1168 the species is confined to the Miocene and ranges from Burdigalian to Tortonian; the species is not recorded at 1139. These ranges fall within the middle-Eocene–late-Miocene (Tortonian) range determined by Holbourn et al. (2013).
Remarks
The distribution is cosmopolitan (Holbourn et al., 2013) and has previously been recorded on the Kerguelen Plateau, southern Indian Ocean, at ODP Sites 748 and 751 (Mackensen, 1992) and 736, 738 and 744 (Schroder-Adams, 1991).
-
Bolivina reticulata Hantken, 1875
-
Figure 3: 3a–b
-
1875 Bolivina reticulata Hantken: p. 65, pl. 15, fig. 6
-
1978 Bolivina reticulata Hantken; Wright, p. 711, pl. 2, figs. 8–10
Description
Broad biserial test with wide, slightly inflated, elongate chambers, characteristically covered by narrow anastomosing ribs. The test is slightly perforated and has a narrow keel around the periphery. Depressed and limbate sutures are observed. The aperture is sub-rounded and located at the base of the last chamber with an internal tooth plate. The species is characterised by its sub-rounded shape in side view and slightly compressed in cross section.
Dimensions
The species length ranged from 150 to 375 µm in the current study.
Stratigraphic range
The species is recorded at 752 from the Chattian to Serravallian, from the Miocene and early Pliocene (Zanclean) at 1168 and Langhian–Tortonian at 1139.
Remarks
This species is recorded in the Mediterranean (Wright, 1978), South Atlantic Ocean (Tjalsma, 1983). This species has a highly variable morphology both in terms of test shape and ornamentation (Wright, 1978). This species is included by some authors as a junior synonym of Latibolivina subreticulata (Parr, 1932) by Loeblich and Tappan (1994) and Holbourn et al. (2013), and it is primarily recorded from the tropical Indian Ocean and Pacific Ocean (Holbourn et al., 2013). Thus, this species likely has a global distribution.
-
Bolivina cf. reticulata Hantken, 1875
-
Figure 3: 5a–b
-
cf. 1875 Bolivina reticulata Hantken: p. 65, pl. 15, fig. 6
-
cf. 1978 Bolivina reticulata Hantken; Wright, p. 711, pl. 2, figs. 8–10
-
cf. 2013 Latibolivina subreticulata (Parr); Holbourn et al., p. 327, figs. 1–4
Description
The test is biserial, with hyaline wall composition. The chamber shape is elongate to trapezoid. The suture is depressed and limbate. The ornamentation is of anastomosing ribs. The aperture is basal and rounded with a tooth plate.
Dimensions
The species length ranges from 125 to 375 µm in the current study.
Stratigraphic range
The species is only observed at 752 through the Miocene and early Pliocene (Zanclean). This species is highly abundant in the mid-to-late Miocene.
Remarks
This species is similar to Bolivina reticulata (Hantken, 1875) but B. reticulata has a broader (particularly towards the base) and relatively more elongate test, with a more compressed periphery in apertural view. According to Wright (1978), Bolivina reticulata varies in shape from an elongate form with limbate sutures to specimens with little ornamentation. This species is also similar to Bolivina viennensis (Marks, 1951) reported in the Polish Carpathian Foredeep Basin (Central Paratethys) by Peryt (2013). This form has so far only recorded in the Indian Ocean.
-
Bolivina sp. 3
-
Figure 3: 4a–c
Description
The test is biserial and elongate with smooth surface. Some specimens are highly perforated. The chambers are elongate to trapezoid in shape. The sutures are slightly depressed. The aperture is sub-rounded and located at the base of the last chamber.
Dimensions
The species length ranged from 150 to 250 µm in the current study.
Stratigraphic range
The species is only observed in the late Oligocene (Chattian) of 752. At 1139 it occurs between the Chattian and Serravallian. At 1168 the species ranges throughout the Miocene and into the early Pliocene (Zanclean).
Remarks
This species differs from Bolivina dilatata (Reuss, 1850), by having a longer test relative to its width (2.5 times long as wide rather than 2:1); the sutures are extremely shallow. The current species is similar to the Bolivina cf. dilatata maxima (p. 148, fig. 7, pl. k) recorded in the Central Paratethys (Peryt, 2013) but the latter has more depressed sutures. The current species differs from Bolivina spathulata (Williamson, 1858), which has sharp edge in the apertural view, whereas specimens here have a sub-rounded edge in apertural view. Also, specimens from the China seas (Lei and Li, 2016) have an anastomosing imperforate costae ornamentation, and they are much larger ∼550 µm (2 times longer than those observed here). This species is so far only been recorded in the southern Indian Ocean (current study).
-
Bolivina sp. 9
-
Figure 3: 6a–b
Description
The test is finely perforated, smooth and elongate, with a hyaline wall composition. Chambers are broad, trapezoid and biserially arranged, with a depressed suture between chambers. Aperture is loop-shaped with basal position. The test length is nearly 3 times longer than the width. Test is tapered at both ends.
Dimensions
The species length ranges from 125 to 250 µm in the current study.
Stratigraphic range
The species occurs through the Miocene and into the early Pliocene (Zanclean) at 752. The species is very abundant in the early Burdigalian at 752. Only very few specimens have been recorded at 1168, mainly in the Chattian and between the Burdigalian to Serravallian with a very few records in the early Pliocene (Zanclean). This species is not recorded at 1139.
Remarks
This species is similar to Bolivinellina pseudopunctata (Alve and Murray, 2001) recorded in intertidal environments from southern England (Ibid; pl. 2, figs. 7–9). The current species has a wider loop-shaped aperture with the last chamber slightly irregular, while B. pseudopunctata has a narrower, slightly compressed aperture and the shape of the chambers are consistently trapezoidal. This species differs from Bolivina spathulata (Williamson, 1858) by having a big and rounded (loop-shaped) aperture on the apertural face rather than a small and narrow loop-shaped aperture. Also, Bolivina spathulata specimens from the China seas (Lei and Li, 2016) have an anastomosing imperforate costae ornamentation, not been observed in the current study.
-
Bolivina viennensis Marks, 1951
-
Figure 3: 7a–b
-
1951 Bolivina viennensis Marks: p. 60, pl. 7, fig. 1
-
1991 Brizalina albatrossi (Cushman); Nomura, p. 53, pl. 1, fig. 3
-
2005 Bolivina viennensis Marks; Popescu and Crihan, p. 380, pl. 1, figs. 14–16
-
2008 Bolivina viennensis Marks; Baldi and Hohenegger, p. 2, fig. 3.a
Description
The test is biserial, elongated. Chambers elongated to trapezoid, sutures depressed. Surface ornamentation reticulate showing an anastomosing pattern. Aperture rounded, terminal. Normal pores are relatively few and concentrated near the margin of the chambers.
Dimensions
The species length ranges from 175 to 575 µm in the current study.
Stratigraphic range
This species is only present at 752 and ranges from the Burdigalian to Messinian, reappearing in the early Pliocene (Zanclean).
Remarks
This species is similar to Bolivina cf. reticulata but the length of Bolivina viennensis is nearly 3 times longer than the width, and the test has a sharp end near the proloculus, while Bolivina cf. reticulata is shorter and is sub-rounded near the proloculus. This species is recorded as Bolivina viennensis in the southern Vienna Basin (Austria) (Baldi and Hohenegger, 2008) and in Romania (Popescu and Crihan, 2005). Similar specimens are recorded as Brizalina albatrossi in the eastern Indian Ocean at Site 754 in the Broken Ridge (Nomura, 1991).
-
Genus Bolivinellina Saidova, 1975
-
Bolivinellina cf. B. translucens (Phleger & Parker, 1951)
-
Figure 3: 8a–c
-
cf. 1951 Bolivina translucens Phleger & Parker: p. 15, pl. 7, figs. 13, 14
-
cf. 1988 Bolivinellina translucens (Phleger & Parker); Loeblich and Tappan: pl. 547, figs. 6, 7
-
cf. 1999 Bolivina translucens (Phleger & Parker); Ohkushi et al., p. 139, pl. 2, figs. 11, 12
Description
Test is elongate, triserial to biserial, narrow and oval in section. Chambers are narrow and moderately inflated; sutures are depressed and oblique. Wall calcareous, hyaline and smooth; a few pores apparent near the sutures. Aperture is basal and slit-to-loop-shaped with a tooth plate at the base of the aperture. Test nearly 3–4 times longer than wide.
Dimensions
The species length ranges from 175 to 475 µm in this study.
Stratigraphic range
The species is only found at Site 752 where it is distributed throughout the succession, with the greatest abundance in the late Miocene (Messinian) and early Pliocene (Zanclean).
Remarks
The chambers appear triserially arranged in the initial part of the test, with inflated chambers which become less inflated later towards the aperture. Bolivinellina translucens is widely distributed in Gulf of Mexico (Phleger and Parker, 1951) and, more locally, has been recorded along the central West Coast of India (Mazumder et al., 2003) and throughout the Indian Ocean (current study).
-
Superfamily Bolivinitoidea Cushman, 1927
-
Family Bolivinitidae Cushman, 1927
-
Genus Abditodentrix Patterson, 1985
-
Abditodentrix pseudothalmanni (Boltovskoy & de Kahn, 1981)
-
Figure 3: 9a–b
-
1981 Bolivinita pseudothalmanni Boltovskoy & de Kahn: p. 44–46, pl. 1, figs. 1–5
-
1985 Abditodentrix asketocomptella Patterson, p. 139, pl. 1, figs. 1–9
-
1988 Abditodentrix pseudothalmanni (Boltovskoy & de Kahn); Loeblich and Tappan, p. 503, pl. 554, figs. 1–5
-
cf. 1991 Brizalina thalmanni (Renz, 1948), Nomura, p. 53, pl. 1, fig. 4
Description
Test elongate, biserial and sagittate. The test is flattened in side view with a distinctive truncate margin giving it a rectangular appearance in section. Chambers are elongated to trapezoid. The sutures are depressed. Aperture is in a basal position and elliptical in shape with tooth plate infolded on one side. The surface is densely covered with elevated reticulate ornamentation.
Dimensions
The species length ranges from 100 to 425 µm in this study.
Stratigraphic range
Occurs in the late Oligocene (Chattian) in all study sites, ranging into the early late Miocene (early Tortonian) in 1139 and the early Pliocene (Zanclean) at 752 and 1168.
Remarks
Most of the observed specimens are highly compressed in cross section and densely ornamented in the initial portion, near the proloculus, with simple reticulation or no ornamentation near the aperture. This species is recorded in the south-west Pacific Ocean (Hayward et al., 2002) and the south-west Atlantic Ocean (Loeblich and Tappan, 1988). It is recorded as Brizalina cf. thalmanni in the eastern Indian Ocean (Nomura, 1991).
-
Superfamily Buliminoidea Jones in Griffith and Henfrey, 1875
-
Family Buliminidae Jones, 1875
-
Genus Bulimina d'Orbigny, 1826
-
Bulimina truncana Gümbel, 1868
-
Figure 3: 10a–b
-
1868 Bulimina truncana Gümbel: p. 727, pl. 2, figs. 77a, b
-
1884 Bulimina rostrata Brady: p. 408, pl. 5, figs. 14, 15
-
1927 Bulimina alazanensis Cushman: p. 161, pl. 25, fig. 4
-
1940 Bulimina bremneri Finlay: p. 455, pl. 64, figs. 84–86
-
1991 Bulimina alazanensis Cushman: Schroder-Adams, p. 624, pl. 1, fig. 5
-
1991 Bulimina truncana Gümbel: Hermelin, p. 65, pl. 1, figs. 10, 16
-
2002 Bulimina truncana Gümbel: Hayward et al., pl. 1, figs. 9, 10
Description
Test is triserial, elongate and sub-triangular in cross section, tapered in outline. Rounded to sub-rounded in the apertural end and pointed in the apical end. The chambers are inflated with obscured sutures. The longitudinal costae occur on all chambers (although the upper portion of the final three chambers is smooth) and join together to form a spinose protrusion from the earliest chambers. The aperture is loop-shaped with internal tooth plate and surrounded by apertural lip. The greatest width of the test is towards the apertural end.
Dimensions
The species length ranges from 125 to 625 µm in the current study.
Stratigraphic range
The species is recorded from the late Oligocene to Pliocene at 752 and 1168, whereas it is confined to the Miocene (Aquitanian to Serravallian) at 1139.
Remarks
Boersma (1986) noted that Bulimina truncana has a highly variable morphology and thus multiple synonyms are based on differences in the number and extent of costae, their height and orientation. Of these, small forms with fewer costae that almost completely cover the final chamber have been often been attributed to Bulimina rostrata, whereas those with more costae, that also tend to be wavy, are assigned to Bulimina alazanensis. Furthermore, B. rostrata also achieves its maximum width towards the middle of the test and is overall sub-circular in cross section compared to B. alazanensis where the maximum test width occurs closer to the apertural end and the test has a more sub-triangular cross section. B. truncana is very similar to B. alazanaensis but was separated out based on having fewer, straighter costae that begin on the first or second chambers rather than the third (Boersma, 1986). Ultimately overlapping species occurrences at the same sites, difficulties in consistently differentiating taxa and missing holotype specimens (B. alazanensis) have led to them being synonymised (Hayward et al., 2019a) in the World Register of Marine Species (WoRMS). Here, tests are usually sub-triangular in cross section, having parallel, regularly formed costae with the largest width of the test towards that apertural end rather than the middle, and hence they are closest to the forms included in B. truncana. This species is considered to be cosmopolitan and has been recorded in Southwest Pacific Ocean (Hayward et al., 2002), Tasman sea, Coral Sea (Boersma, 1986) and Indian Ocean (Nomura, 1995).
-
Bulimina cf. B. elongata d'Orbigny, 1846
-
Figure 3: 11a–b
-
cf. 1846 Bulimina elongata d'Orbigny: p. 187, pl. 11, figs. 19, 20
-
cf. 1826 Bulimina elongata d'Orbigny: (Nomen nudum), p. 269
-
cf. 2004 Bulimina elongata d'Orbigny: De Man et al., p. 187 , pl. 2, fig. 9
-
cf. 2008 Bulimina elongata d'Orbigny: Kender et al., p. 513, pl. 17, figs. 4, 5
Description
Elongate, narrow (3 times as long as wide), triserial test becoming nearly biserial in final whorl. Little tapering of test. Chambers are flattened and broad. Sutures are depressed. The aperture is basal and rounded. The surface is mainly smooth with very few perforations.
Dimensions
The species length ranges from 150 to 400 µm in this study.
Stratigraphic range
This species ranges from the late Oligocene (Chattian) to mid-Burdigalian at 1168, extending into the early Tortonian at 1168. This supports the range (mid-Eocene to Pleistocene) identified by Thomas (1990). This species is not present at 752.
Remarks
This species differs from Bulimina elongata sensu stricto (see Holbourn et al., 2013) in that the chambers are less inflated (have a nearly polygonal outline) and a much slower rate of chamber enlargement. Holbourn et al. (2013) consider Bulimina elongata a cosmopolitan species.
-
Bulimina gibba Fornasini, 1902
-
Figure 3: 12
-
1902 Bulimina gibba Fornasini: p. 378, pl. O, figs. 32–34
-
2005 Bulimina gibba Fornasini; Rasmussen, p. 86, pl. 11, fig. 5
-
2013 Bulimina gibba Fornasini; Holbourn et al., p. 97, figs. 1–3
Description
Triserial, elongate, fusiform shape. Nearly circular in cross section with narrow apical tip. Chambers are inflated with clearly depressed sutures and strongly overlapping. The aperture is loop-shaped, located at the base of the last chamber with apertural lip merging with an internal tooth plate. The test surface is relatively smooth with perforation distributed across the chambers.
Dimensions
The species length ranges from 125 to 300 µm in the current study.
Stratigraphic range
This species occurs from the late Oligocene (Chattian) to early Tortonian at 1139. This species is not recorded at either 752 or 1168.
Remarks
This species is commonly found and described (e.g. Holbourn et al., 2013) as having an apical protrusion from the proloculus; however, in this study, no spinose protrusions are observed in any specimen. This absence cannot be explained by preservation. This species has previously only been reported from the Atlantic (Holbourn et al., 2013), but the authors noted that it may well range further, which is confirmed in this study. Holbourn et al. (2013) report this species from the Miocene to Holocene but this work suggests it may occur earlier in the Indian Ocean.
-
Bulimina striata d'Orbigny in Guérin-Méneville, 1832
-
Figure 3: 13
-
1832 Bulimina striata d'Orbigny in Guérin-Méneville: p. 18, pl. 3, fig. 16
-
1852 Bulimina costata d'Orbigny: p. 194
-
1922 Bulimina inflata var. mexicana Cushman: p. 95, pl. 21, fig. 2
-
1953 Bulimina mexicana Cushman; Phleger and others, p. 33, pl. 6, fig. 27
-
1954 Bulimina striata mexicana Cushman; Parker, p. 511, pl. 6, fig. 24
-
1957 Bulimina australis Vella: p. 32, pl. 8, figs. 168–169
-
1992 Bulimina striata d'Orbigny in Guérin-Méneville: Mackensen, p. 668, pl. 1, fig. 5
-
2003 Bulimina striata d'Orbigny in Guérin-Méneville: Hayward et al., p. 517
-
2011 Bulimina striata d'Orbigny in Guérin-Méneville: Kuppusamy et al., p. 46, pl. 2, figs. 7, 8
-
2013 Bulimina striata d'Orbigny in Guérin-Méneville: Mancin et al., p. 64, pl. 2, fig. 3
Description
Test is conical, triserial, and semicircular in cross section; the initial part of the test (near proloculus) is acute. Chambers are inflated overhanging previous chambers, becoming broader and less ornamented near the aperture. The sutures are depressed but largely obscured by ornamentation in the earlier portion of the test. Longitudinal costae extend from the lower half of the chambers to overhang preceding chambers ending in acute points. The aperture is loop-shaped and bordered by a lip with an internal tooth plate.
Dimensions
The species length ranges from 100 to 500 µm in this study.
Stratigraphic range
The species is recorded throughout the whole succession (late Oligocene–Pliocene) at 752. At 1139 and 1168 this species occurs from the late Oligocene (Chattian) disappearing from 1139 in the Langhian and early Tortonian at 1168.
Remarks
This species is cosmopolitan. This species is identified around New Zealand (Debenay, 2012; Hayward et al., 2003) and by Kuppusamy et al. (2011) in north-west Atlantic Ocean. Many older Indian Ocean publications report this species as B. mexicana (e.g. Nomura, 1995), which does have a global distribution (Holbourn et al., 2013). Holbourn et al. (2013) consider the range of this species to be Miocene–Holocene; here it is found in the late Oligocene.
-
Superfamily Turrilinoidea Cushman, 1927
-
Family Turrilinidae Cushman, 1927
-
Subfamily Turrilininae Cushman, 1927
-
Genus Eubuliminella Revets, 1993
-
Eubuliminella exilis (Brady, 1884)
-
Figure 4: 1
-
1884 Bulimina elegans var. exilis Brady: p. 399, pl. 50, figs. 5, 6
-
1927 Buliminella subfusiformis var. tenuata Cushman: p. 149, pl. 2, fig. 9
-
1930 Buliminella subfusiformis (Cushman); Cushman and Moyer, p. 56, pl. 7, fig. 20
-
1947 Bulimina exilis Brady; Cushman and Parker, p. 123, pl. 28, figs. 27, 28
-
1947 Bulimina subfusiformis var. tenuata (Cushman); Cushman and Parker, p. 124, pl. 28, fig. 29
-
1993 Eubuliminella exilis (Brady); Revets, p. 141, pl. 1, figs. 1–7
-
2015 Eubuliminella exilis (Brady); Setoyama & Kaminski, p. 8, figs. 4.2, 8.7
Description
Elongate, narrow test that is triserial in the latter part. The test length is about 4 times as long as it is wide. Ovate and little inflated chambers separated by curved and depressed sutures. The aperture is basal and loop-shaped, but from the apertural opening the tooth plate is not clear.
Dimensions
The species length ranges from 150 to 550 µm in the current study.
Stratigraphic range
The stratigraphic range of the species at 1168 is early Miocene (Burdigalian) to the early Pliocene (Zanclean); Holbourn et al. (2013) suggested a range from the late Miocene; the present study therefore extends the known stratigraphic range.
Remarks
The specimens in the current study lack the spine in the initial whorl and have more weakly inflated chambers. According to Revets (1993) and Holbourn et al. (2013), this species can be found worldwide.
-
Superfamily Cassidulinoidea d'Orbigny, 1839a
-
Family Cassidulinidae D'Orbigny, 1839a
-
Subfamily Cassidulininae d'Orbigny, 1839a
-
Genus Globocassidulina Voloshinova, 1960
-
Globocassidulina subglobosa (Brady, 1881)
-
Figure 4: 2
-
1881 Cassidulina subglobosa Brady: p. 60, illustrated in Brady, 1884, pl. 54, fig. 17
-
1884 Cassidulina subglobosa Brady; Brady, p. 430, pl. 54, fig. 17
-
1983 Globocassidulina subglobosa (Brady); Tjalsma and Lohmann, p. 31, pl. 16, fig. 9
-
1991 Globocassidulina subglobosa (Brady); Nomura, p. 55, pl. 5, fig. 8
-
2013 Globocassidulina subglobosa (Brady); Holbourn et al., p. 264, figs. 1, 2
Description
Test is biserial and sub-globular, with a sub-circular shape in cross section. Chambers are inflated and separated by depressed, curved sutures. The test wall is smooth and only finely perforate. The aperture is slit-like, straight or curved.
Dimensions
The species length ranges from 100 to 925 µm in this study.
Stratigraphic range
This is a long-ranging species, recorded from late Oligocene to early Pliocene at both 752 and 1168 and late Oligocene to the top of the studied interval (early Tortonian) at 1139.
Remarks
This species is very well-preserved in all studied sites. This is a cosmopolitan species (Jones, 1994) that is very long-ranging (Paleocene–Holocene; Tjalsma and Lohmann, 1983).
-
Genus Paracassidulina Nomura, 1983b
-
Paracassidulina minuta (Cushman, 1933b)
-
Figure 4: 3a–b
-
1933b Cassidulina minuta Cushman: p. 92, pl. 10, figs. 3a–c
-
1951 Cassidulina crassa d'Orbigny; Phleger and Parker, p. 26, pl. 14, figs. 4a–b
-
1954 Cassidulina aff. crassa d'Orbigny; Parker, p. 535, pl. 11, fig. 1
-
1983b Paracassidulina minuta Cushman; Nomura, p. 66, pl. 5, figs. 16a–c
-
1991 Paracassidulina minuta Cushman; Nomura, p. 55, pl. 5, figs. 2a–b
Description
Biserial test with slightly depressed sutures. Surface is smooth and finely perforated. Chambers are strongly curved. Aperture is long, curved and has an apertural ridge parallel to the chamber periphery.
Dimensions
The species length ranges from 125 to 300 µm in the current study.
Stratigraphic range
The species ranges from Chattian to early Tortonian at 752, Chattian to mid-Serravallian at 1139. The stratigraphic distribution of the species at 1168 is early Aquitanian to Zanclean.
Remarks
The chambers of the specimens found here are only slightly inflated, this differs from Globocassidulina minuta (Cushman, 1933b) described from the Paumotu Islands (also called Tuamotu Islands) in the Southern Pacific Ocean in that the latter has more globular chambers. This species is recorded by Nomura (1991) at Broken Ridge (Site 754) in eastern Indian Ocean and in the Gulf of Mexico (Poag, 2015).
-
Subfamily Ehrenbergininae Cushman, 1927
-
Genus Ehrenbergina Reuss, 1850
-
Ehrenbergina carinata Eade, 1967
-
Figure 4: 4a–b
-
1967 Ehrenbergina carinata Eade: p. 448, pl. 9, figs. 1–4
-
1991 Ehrenbergina carinata Eade; Nomura, p. 55, pl. 5, fig. 13
-
2013 Ehrenbergina carinata Eade; Hayward et al., p. 442, pl. 6, figs. 12, 13
Description
Test triangular in both dorsal view and cross section. Early chambers are not inflated, while the later chambers are slightly inflated. Well-developed peripheral keel and a spine extending from each chamber. Dorsal view is compressed. The sutures are flush but towards the periphery become raised. The ventral view is characterised by central keel. Aperture is slit-like and curved, with a narrow lip in the ventral side.
Dimensions
The species length ranges from 150 to 725 µm in the current study.
Stratigraphic range
The species range at 752 is late Miocene (Messinian) to early Zanclean. At 1168 the species is very rare and only recorded in the early and late Miocene (Tortonian). This species is not recorded at 1139.
Remarks
This species is well-preserved at 752 and reaches very large size (up to 725 µm) in some intervals, especially in the late Miocene (Messinian). According to Eade (1967) the current species is recorded in the Kermadec Islands, Fiji, Norfolk Island, Tonga, and northern New Zealand. This species is also recorded by Nomura (1991) at Broken Ridge (Site 754) and New Zealand (Hayward et al., 2013).
-
Genus Burseolina Seguenza, 1880
-
Burseolina pacifica (Cushman, 1925)
-
Figure 4: 5a–b
-
1925 Cassidulina pacifica Cushman: p. 53, figs. 14–16
-
1925 Globocassidulina pacifica (Cushman), pl. 2, fig. 16
-
1983a Burseolina pacifica (Cushman); Nomura, pl. 6, fig. 2, pl. 21, figs. 6–10
-
1983b Burseolina pacifica (Cushman); Nomura, pp. 57–60, pl. 5, figs. 1–4
-
1991 Burseolina cf. pacifica (Cushman); Nomura, p. 53, pl. 5, figs. 14a–b
Description
Enrolled biserial test with rounded periphery. Sutures are flush to the surface and thus almost indistinguishable. The apertural face is flattened and the aperture is elongate, curved, and with apertural flap in the lower apertural margin.
Dimensions
The species length ranges from 175 to 600 µm in this study.
Stratigraphic range
The distribution of the species at 752 ranges from the late Oligocene (Chattian), where it is rare, with later Miocene records from the Burdigalian to mid-Messinian. The species is rare at 1168 in the Chattian and only appears in the late Burdigalian in the Miocene. This species is not recorded at 1139.
Remarks
According to Cushman (1925) this species is widely distributed in deep or cool waters and it has been recorded in the Pacific Ocean. It has also been reported from subtropical regions, along the Pacific coast of Japan (Nomura, 1983a) and in the Indian Ocean (Nomura, 1991). Notably the specimens here are more spherical in outline than B. pacifica reported by Nomura (1991). Burseolina is most abundant from the Miocene to the present.
-
Burseolina cf. pacifica (Cushman, 1925)
-
Figure 4: 6
-
cf. 1925 Cassidulina pacifica Cushman: p. 53, figs. 14–16
-
cf. 1925 Globocassidulina pacifica (Cushman), pl. 2, fig. 16
-
cf. 1983a Burseolina pacifica (Cushman); Nomura, pl. 6, fig. 2, pl. 21, figs. 6–10
-
cf. 1983b Burseolina pacifica (Cushman); Nomura, pp. 57–60, pl. 5, figs. 1–4
-
cf. 1991 Burseolina pacifica (Cushman); Nomura, p. 53, pl. 5, figs. 4a–b
Description
Test is biserial and sub-spherical. The surface is smooth. Flush to slightly depressed sutures. The aperture is elongated, curved and with apertural flap in the lower apertural margin.
Dimensions
The species length ranges from 100 to 425 µm in this study.
Stratigraphic range
The range of the species at 752 is early Miocene (Aquitanian) to early Pliocene (Zanclean). At 1139 it is very rare in the late Oligocene (Chattian) and also in the Miocene, except for Serravallian where it has its greatest abundance. The distribution at 1168 is mid-Burdigalian to Zanclean.
Remarks
This species differs from Burseolina pacifica by having a relatively larger and wider aperture and broader flap. The last chambers in this species are also usually separated from the preceding chambers by clearly depressed sutures. This species has a limited geographic distribution and is also observed in tropical to subtropical regions, along the Pacific coast of Japan (Nomura, 1983a) and in the Indian Ocean on the Ninetyeast Ridge (Site 756) (Nomura, 1991).
-
Superfamily Pleurostomelloidea Reuss, 1860
-
Family Pleurostomellidae Reuss, 1860
-
Subfamily Pleurostomellinae Reuss, 1860
-
Genus Pleurostomella Reuss, 1860
-
Pleurostomella acuminata Cushman, 1922
-
Figure 4: 7a–b
-
1922 Pleurostomella acuminata Cushman: p. 50, pl. 19, fig. 6
-
1866 Pleurostomella alternans Schwager: p. 238, pl. 6, fig. 80
-
1934 Pleurostomella alternans var. telostoma Schubert; Cushman, pl. 16, fig. 6
-
1978 Pleurostomella acuta Hantken; Boltovskoy, pl. 5, fig. 42
-
1978 Pleurostomella dominicana Bermúdez; Boltovskoy, pl. 6, figs. 2, 3
-
2011 Pleurostomella acuminata (Cushman); Kuppusamy et al., p. 60, pl. 9, figs. 7, 8
Description
Test is smooth, elongate and biserial, with slightly depressed sutures. Chambers are elongate and oval. Wide terminal aperture, with projecting hood on one side and two triangular or bifid teeth on opposite side. Prolocular spine is present.
Dimensions
The species length ranges from 150 to 725 µm in the current study.
Stratigraphic range
At 752 and 1168 it ranges from Chattian to early Zanclean and at 1139 from Chattian to early Tortonian.
Remarks
A prolocular spine is present in most specimens but in others it is either broken or missing. This species is cosmopolitan and recorded throughout the Indian Ocean (Hayward, 2002).
-
Superfamily Stilostomelloidea Finlay, 1947
-
Family Stilostomellidae Finlay, 1947
-
Genus Orthomorphina Stainforth, 1952
-
Orthomorphina perversa (Schwager, 1866)
-
Figure 4: 8a–c
-
1866 Nodosaria perversa Schwager: p. 212, pl. 5, fig. 29
-
1872 Nodosaria pupoides Silvestri: p. 65, pl. 6, figs. 148–158
-
1937 Nodogenerina challengeriana Thalmann: p. 341, pl. 64, figs. 25–27
-
1949 Nodogenerina aminaensis Bermúdez: p. 178, pl. 11, fig. 57
-
1978 Orthomorphina challengeriana (Thalmann); Boltovskoy, pl. 5, figs. 16–17
-
1978 Orthomorphina perversa (Schwager); Boltovskoy, pl. 5, figs. 23–24
-
1992 Stilostomella sp. G. Kaiho: p. 307, pl. 5, fig. 23
-
2002 Orthomorphina perversa (Schwager); Hayward, pl. 1, figs. 50–52
Description
Test uniserial, rectilinear. Chambers are ovoid in outline and increase in height towards the last chamber. The surface is striate with ribs perpendicularly arranged. The sutures are straight and depressed. The aperture is rounded and terminal with rim.
Dimensions
The species length ranges from 200 to 975 µm in the current study.
Stratigraphic range
The species ranges from Chattian to early Zanclean at 752 and 1168 but only extends into the mid-Serravallian (mid-Miocene) at 1139.
Remarks
The largest chamber is usually in the middle part of the test and the last chamber is smaller than the penultimate one. Sometimes this difference is very pronounced. This species is cosmopolitan (Hayward, 2002).
-
Genus Siphonodosaria Silvestri, 1924
-
Siphonodosaria lepidula Schwager, 1866
-
Figure 4: 9a–b
-
1866 Nodosaria lepidula Schwager: p. 210, pl. 5, figs. 27, 28
-
1978 Stilostomella ex.gr. lepidula (Schwager). Boltovskoy, pl. 7, figs. 33–36
-
1980 Stilostomella lepidula (Schwager); Srinivasan and Sharma, p. 46, pl. 7, figs. 1–6
-
1993 Stilostomella lepidula (Schwager); Gupta, 1993, figs. 2, 1–2
-
1994 Nodogenerina lepidula (Schwager); Loeblich and Tappan, p. 133, pl. 261, figs. 18–20
-
2002 Siphonodosaria lepidula f. lepidula (Schwager); Hayward, p. 305, pl. 3, figs. 25–32
-
2012 Siphonodosaria lepidula (Schwager); Hayward et al., p. 174, pl. 18, figs. 6–20
Description
Elongate, uniserial test, slightly arcuate shape, with chambers gradually increasing in size. Chambers are spherical (campanulate or pyriform) and typically become more widely spaced later with pronounced depressed sutures. A ring of small spines, annularly arranged, surrounds the base of each otherwise relatively smooth-walled chamber. Aperture is terminal with a broad neck and phialine lip; there is small tooth in the middle of the aperture giving a v-shaped appearance to aperture.
Dimensions
The species length ranges from 200 to 1350 µm in this study.
Stratigraphic range
The distribution at 752 and 1168 is Chattian to early Zanclean and at 1139 is Chattian to early Tortonian.
Remarks
This species is abundant, with a cosmopolitan distribution (Hayward et al., 2012; Holbourn et al., 2013). It is also highly morphologically variable showing considerable variation in ornamentation, chamber shape and the presence or absence of a prolocular spine, and hence has been frequently recorded under a number of different names (Hayward et al., 2012). After detailed population level investigations, Hayward et al. (2012) group the different morphotypes under S. lepidula is known from the Late Cretaceous to Pleistocene, reports of recent occurrences are considered reworked (Hayward et al., 2012).
-
Siphonodosaria subspinosa (Cushman, 1943)
-
Figure 4: 10a–d
-
1934 Ellipsonodosaria sp. Cushman and Jarvis: pl. 10, figs. 4, 5
-
1943 Ellipsonodosaria subspinosa Cushman: p. 92, pl. 16, figs. 6, 7b
-
1983 Stilostomella subspinosa (Cushman); Tjalsma and Lohmann, p. 36, pl. 14, figs. 16, 17
-
1998 Siphonodosaria subspinosa (Cushman); Robertson, p. 180, pl. 67, fig. 3
-
2010 Siphonodosaria subspinosa (Cushman); Hayward et al., p. 129, pl. 14, figs. 20, 21
Description
Test elongate, uniserial, straight to slightly arcuate shape, circular in cross section. Chambers are spherical, separated by strongly depressed sutures. Short spines cover the surface. Chambers show a relatively rapid increase in size towards the aperture creating a tapered test outline. The aperture is terminal with broad neck and phialine lip.
Dimensions
The species length ranges from 200 to 1125 µm in the current study.
Stratigraphic range
The distribution at 752 and 1168 is Chattian to early Zanclean, and at 1139 Chattian to early Tortonian.
Remarks
Specimens are very well-preserved and abundant at Site 752. Holbourn et al. (2013) comment that this species shows large variability in the size and ornamentation of the test, with short spines commonly found on the test sometimes converging to form fine costae in the lower portion of the test. This is consistent with observations in this study. Distribution is worldwide (Holbourn et al., 2013).
-
Genus Stilostomella Guppy, 1894
-
Stilostomella cf. S. annulifera Cushman and Bermúdez, 1936
-
Figure 4: 11a–b
-
cf. 1936 Ellipsonodosaria annulifera Cushman and Bermúdez: p. 28, pl. 5, figs. 8, 9
-
1978 Stilostomella cf. S. annulifera Wright: p. 717, pl. 8, fig. 2
-
cf. 1990 Stilostomella annulifera (Cushman and Bermúdez); Thomas, 1990, p. 590, pl. 1, fig. 4
-
cf. 1998 Siphonodosaria annulifera (Cushman and Bermúdez); Robertson, p. 178, pl. 66, figs. 1, 2
Description
Test elongate, rectilinear, arcuate in overall shape with a smooth wall texture. Chambers are inflated to sub-spherical. Sutures are limbate and shallow. Chambers become bigger and more elongated towards apertural end. Aperture is terminal with phialine lip.
Dimensions
The species length ranges from 225 to 1250 µm in this study.
Stratigraphic range
The distribution of the species at 1139 is Chattian to early Tortonian and at 1168 and 752 from Chattian to early Zanclean.
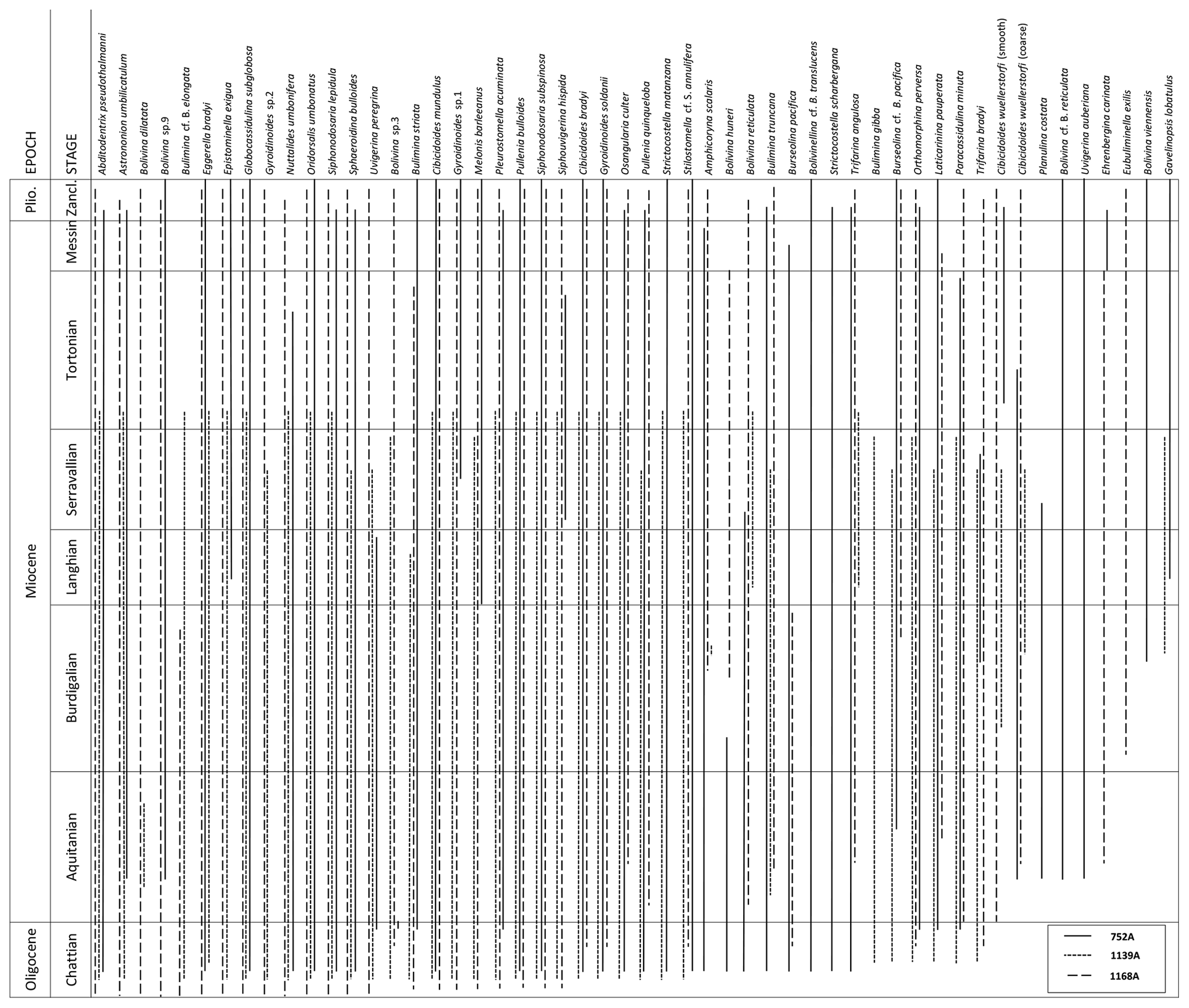
Figure 2Range chart of the 52 most abundant benthic foraminifera species encountered at Ocean Drilling Program (ODP) Holes 752A, 1139A and 1168A.

Figure 3Scale bar is 100 µm. (1a–b) Bolivina dilatata, ODP Hole 1168A, 37X-6, 75–79 cm: (1a) side view; (1b) apertural view. (2a–b) Bolivina huneri, ODP Hole 752A, 10H-5, 77–81 cm. (2a) side view; (2b) apertural view. (3a–b) Bolivina reticulata, ODP Hole 1168A, 15X-2, 76–80 cm: (3a) side view; (3b) apertural view. (4a–b) Bolivina sp. 3, ODP Hole 1168A, (4a–b) 26X-2, 73–77 cm: (4a) side view; (4b) apertural view. (4c) 37X-6, 75–79 cm: side view. (5a–b) Bolivina cf. reticulata, ODP Hole 752A, (5a) 5H-1, 65–70 cm: apertural view. (5b) 3H-5, 62–67 cm: side view. (6a–b) Bolivina sp. 9, ODP Hole 752A, 9H-2, 104–109 cm: (6a) side view; (6b) apertural view. (7a–b) Bolivina viennensis, ODP Hole 752A, 5H-1, 65–70 cm: (7a) side view; (7b) apertural view. (8a–c) Bolivinellina cf. B. translucens, ODP Hole 752A, 3H-1, 50–55 cm: (8a–b) side view; (8c) apertural view. (9a–b) Abditodentrix pseudothalmanni, ODP Hole 1139A, 19R-2, 60.5–64.5 cm: (9a) side view; (9b) apertural view. (10a–b) Bulimina truncana, (10a) ODP Hole 752A, 9H-5, 65–70 cm: lateral view. (10b) ODP Hole 1168A, 37X-6, 75–79 cm: apertural side view. (11a–b) Bulimina cf. B. elongata, (11a) ODP Hole 1168A, 23X-3, 74–78 cm: apertural side view. (11b) ODP Hole 1139A, 18R-1, 96–100 cm: apertural side view. (12) Bulimina gibba, ODP Hole 1139A, 19R-2, 60.5–64.5 cm: lateral view. (13) Bulimina striata, ODP Hole 752A, 5H-1, 65–70 cm: apertural side view.
Remarks
Specimens here largely lack the prolocular spine because of preservation (see Fig. 4: 11a). This species is similar to Strictocostella scharbergana Neugeboren, 1856, but the last chamber of S. annulifera is slightly bigger and more inflated with a shorter neck. Some publications (e.g. Hayward et al., 2012) synonymise these two species and use S. scharbergana. S. consobrina differs from this species by having sutures which are flusher to the test, as shown in Holbourn et al. (2013). It has also been observed that the chambers of this species are particularly large in those specimens recorded in Mediterranean Sea (Wright, 1978). S. cf. S. annulifera has a cosmopolitan distribution (Holbourn et al., 2013) including records in the Mediterranean Sea (Wright, 1978) and Antarctica (Thomas, 1990). S. annulifera ranges from the early Eocene to middle Miocene (Holbourn et al., 2013), but S. scharbergana is considered to range from the early Paleocene to mid-Pleistocene (Hayward et al., 2012), more consistent with this extended younger occurrence.
-
Genus Strictocostella Patterson, 1987
-
Strictocostella matanzana (Palmer & Bermúdez, 1936)
-
Figure 4: 12a–c
-
1936 Ellipsonodosaria? matanzana Palmer & Bermúdez: p. 298, pl. 18, fig. 12
-
1940 Ellipsonodosaria matanzana Palmer and Bermúdez; Palmer, pl. 15, fig. 1
-
1957 Siphonodosaria cf. matanzana (Palmer and Bermúdez); Todd, pl. 67, fig. 7
-
1975 Stilostomella lepidula Saidova: p. 315, pl. 87, fig. 6
-
1998 Siphonodosaria matanzana (Palmer and Bermúdez); Robertson, pl. 67, fig. 2
-
2007 Myllostomella matanzana (Palmer and Bermúdez); O'Neill et al., p. 1085, pl. 1, figs. 22–23
-
2010 Strictocostella matanzana (Palmer and Bermúdez); Hayward et al., p. 128, figs. 14, 27–28
-
2012 Strictocostella matanzana (Palmer and Bermúdez); Hayward et al., p. 190, pl. 22, figs. 10–20

Figure 4Scale bar is 100 µm. (1) Eubuliminella exilis, ODP Hole 1168A, 42X-3, 70–74 cm: apertural side view. (2) Globocassidulina subglobosa, ODP Hole 752A, 4H-1, 70–75 cm: apertural side view. (3a–b) Paracassidulina minuta, ODP Hole 1168A, (3a) 42X-3, 70–74 cm: apertural side view. (3b) 37X-6, 75–79, 0–1 cm: side view. (4a–b) Ehrenbergina carinata, ODP Hole 752A, 3H-1, 50–55 cm: (4a) apertural side view; (4b) side view. (5a–b) Burseolina pacifica, ODP Hole 752A, 6H-1, 70–75 cm: (5a) apertural side view; (5b) side view. (6) Burseolina cf. pacifica, ODP Hole 752A, 6H-1, 70–75 cm: apertural side view. (7a–b) Pleurostomella acuminata, (7a) ODP Hole 752A, 7H-1, 63–68 cm: apertural side view. (7b) ODP Hole 1168A, 22X-2, 45–49 cm: lateral view. (8a–c) Orthomorphina perversa, (8a) ODP Hole 1139A, 14R-4, 65–69 cm: side view. (8b, c) ODP Hole 752A, 3H-1, 50–55 cm: (8b) side view; (8c) apertural view. (9a–b) Siphonodosaria lepidula, ODP Hole 752A, (9a) 7H-2, 73–78 cm: side view. (9b) ODP Hole 8H-1, 100–105 cm: apertural view. (10a–d) Siphonodosaria subspinosa, (10a–b) ODP Hole 752A, 8H-6, 70–75 cm: (10a) side view; (10b) apertural view. (10c) ODP Hole 1168A, 18X-5, 80–84 cm: side view. (10d) ODP Hole 1168A, 15X-2, 76–80 cm: apertural view. (11a–b) Stilostomella cf. S. annulifera, ODP Hole 1139A, 15R-4, 41–45 cm: (11a) side view; (11b) apertural view. (12a–c) Strictocostella matanzana, (12a–b) ODP Hole 752A, 3H-5, 62–67 cm: (12a) side view; (12b) apertural view. (12c) ODP Hole 1139A, 18R-1, 96–100 cm: side view.
Description
Test elongate, uniserial, rectilinear. The base is composed of few small, spherical chambers which become larger and more elongate towards the aperture. Sutures are straight and depressed. There are small spines forming a ring towards the base of each chamber. The aperture is terminal with a phialine lip and short, broad neck ornamented by pustular spines.
Dimensions
The species length ranges from 200 to 750 µm in the current study.
Stratigraphic range
The species occurs rarely in the Chattian at Site 752 and it is not recorded in Aquitanian–Langhian, but appears again from early Serravallian to Zanclean. At 1139 the distribution of the species is from Chattian to early Tortonian; it is not recorded at 1168.

Figure 5Scale bar is 100 µm. (1a–b) Strictocostella scharbergana, ODP Hole 752A, 4H-1, 70–75 cm: (1a) side view; (1b) apertural view. (2a–c) Trifarina angulosa, (2a) ODP Hole 752A, 6H-1, 70–75 cm: side view. (2b, c) ODP Hole 1168A, 23X-3, 74–78 cm: (2b) side view; (2c) apertural view. (3a–c) Trifarina bradyi, (3a) ODP Hole 752A, 8H-6, 70–75 cm: side view. (3b) ODP Hole 1168A, 15X-2, 76–80 cm: side view. (3c) ODP Hole 1139A, 5R-1, 121–125 cm: side view. (4a–c) Siphouvigerina hispida, (4a–b) ODP Hole 1168A, 20X-3, 100–104 cm: (4a) side view; (4b) apertural view. (4c) ODP Hole 1139A, 7R-1, 111–115 cm: side view. (5a–d) Uvigerina auberiana, ODP Hole 752A, (5a) 10H-2, 60–65 cm: side view. (5b) 3H-5, 62–67 cm: side view. (5c, d) 4H-2, 81–85 cm: (5c) side view; (5d) apertural view. (6a–b) Uvigerina peregrina, (6a) ODP Hole 752A, 9H-5, 65–70 cm: side view. (6b) ODP Hole 1168A, 22X-2, 45–49 cm: side view. (7a–b) Lenticulina cultrata, ODP Hole 752A, 10H-5, 77–81 cm; (7a) side view; (7b) edge view.
Remarks
The wall of the neck is folded, giving an undulation to the phialine lip. S. matanzana has a cosmopolitan distribution; sister species tend to be more prevalent in the Indian Ocean (Hayward et al., 2012).
-
Strictocostella scharbergana (Neugeboren, 1856)
-
Figure 5: 1a–b
-
1856 Dentalina scharbergana Neugeboren: p. 87, pl. 4, figs. 1–4
-
1856 Strictocostella scharbergana Neugeboren, p. 87, pl. 4, figs. 1–4
-
1866 Dentalina costai Schwager: p. 229, pl. 6, fig. 62
-
1884 Nodosaria consobrina (d'Orbigny); Brady, pl. 62, figs. 23, 24
-
1936 Ellipsonodosaria annulifera Cushman and Bermúdez: p. 28, pl. 5, figs. 8, 9
-
1960 Stilostomella consobrina (d'Orbigny); Barker, pl. 62, figs. 23, 24
-
1977 Siphonodosaria consobrina (d'Orbigny); Nishimura et al., pl. 4, figs. 16, 17
-
1994 Stilostomella consobrina (d'Orbigny); Jones, 1994, pl. 62, figs. 23, 24
-
2000 Stilostomella scharbergana (Neugeboren); Sztràkos, pl. 5, fig. 19
-
2005 Myllostomella costai (Schwager); Hayward and Kawagata, pl. 1, figs. 17, 18
-
2007 Strictocostella costai (Schwager); Hayward et al., fig. 2
-
2010 Strictocostella scharbergana (Neugeboren); Hayward et al., p. 128, figs. 14, 28–29
-
2012 Strictocostella scharbergana (Neugeboren); Hayward et al., p. 192, pl. 23, figs. 8–19
Description
Test arcuate, uniserial, slender. Proloculus spherical but chambers become more ovate towards the aperture. Surface is smooth. Sutures flush or slightly incised. Aperture is terminal with a short neck and phialine lip, internal apertural rim consists of a long, simple tooth and multiple long, fine denticles.
Dimensions
The species length ranges from 200 to 1400 µm in the current study.
Stratigraphic range
The species range at 752A is Chattian to early Zanclean. This species is not recorded at 1168 and 1139.
Remarks
Wall smooth throughout, occasionally with one pustular spine near the base of the proloculus. Hayward et al. (2012) consider S. scharbergana conspecific with Stilostomella consobrina, and in many works it is identified as S. scharbergana (e.g. Sztràkos, 2000; Hayward et al., 2010, 2012). Cosmopolitan distribution (Hayward et al., 2012).
-
Family Uvigerinidae Haeckel, 1894
-
Subfamily Angulogerininae Galloway, 1933
-
Genus Trifarina Cushman, 1923
-
Trifarina angulosa (Williamson, 1858)
-
Figure 5: 2a–c
-
1858 Uvigerina angulosa Williamson: p. 67, pl. 5, fig. 140
-
1960 Angulogerina angulosa (Williamson); Barker, pl. 74, figs. 15, 16
-
1994 Trifarina angulosa (Williamson); Jones, p. 86, pl. 74, figs. 15, 16
-
2013 Trifarina angulosa (Williamson); Holbourn et al., p. 559, figs. 1–4
Description
The test is triserial in the early portion, becoming uniserial later. Test cross section is triangular with acute periphery. The chambers are moderately inflated and increase in size towards the apertural end. The sutures are curved and depressed. Wall ornamented by discontinuous longitudinal costae. The aperture is terminal, with a rounded opening atop a short neck, with lip and internal tooth plate.
Dimensions
The species length ranges from 150 to 775 µm in the current study.
Stratigraphic range
This species shows a staggered appearance; first appearing sporadically at Site 752 in the Chattian then in the Aquitanian at 1168, followed by the mid-Langhian at 1139. The last appearance is in the Tortonian at 1139 and in the Zanclean at 1168 and 752.
Remarks
Specimens from 1168 usually have the typical triangular cross section with acute periphery but those from 1139 and particularly 752 often appear nearly rounded in cross section and lack the acute margin. This species is typically recorded from the mid-Miocene to Holocene (Jones, 1994). Thus, this study extends the species range back into the latest Oligocene. The species is found worldwide (Holbourn et al., 2013).
-
Trifarina bradyi Cushman, 1923
-
Figure 5: 3a–c
-
1923 Trifarina bradyi Cushman: p. 99, pl. 22, figs. 3, 9
-
1884 Rhabdogonium tricarinatum d'Orbigny; Brady, p. 525, pl. 67, figs. 1, 3
-
1960 Trifarina bradyi Cushman; Barker, pl. 67, figs. 1, 3
-
1988 Trifarina bradyi Cushman; Loeblich and Tappan, p. 526, pl. 574, figs. 10–13
-
2013 Trifarina bradyi Cushman; Holbourn et al., p. 560, figs. 1, 2
Description
Test initially triserial becoming rectilinear and uniserial later. Test tapered at both ends and relatively smooth, later chambers. Distinctive triangular cross section with acute peripheral keel. Chambers are moderately inflated and increase in size towards the aperture, separated by thick, strongly curved, depressed sutures. The aperture is terminal with short neck, lip and tooth plate.
Dimensions
The species length ranges from 125 to 575 µm in this study.
Stratigraphic range
At 1139 and 1168 the species is rare in the Chattian, but is more consistently present from the Aquitanian disappearing in the mid-Serravallian at 1139 and early Zanclean at 1169. The species at 752 is restricted to the Burdigalian through to Serravallian (mid-Miocene).
Remarks
Inter-site differences are observed within this species in the Indian Ocean. Specimens from 752 and 1139 are quite similar, with the lower half of the test having longitudinal striations, while at 1168 there are denticles or small nodes (see Fig. 5: 3a and b). Distribution is worldwide according to Holbourn et al. (2013) and it has previously been recorded from the eastern Indian Ocean by Nomura (1991). As with T. angulosa, a much earlier late Oligocene occurrence for this species is found in the eastern Indian Ocean than previously reported (Jones, 1994).
-
Subfamily Uvigerininae Haeckel, 1894
-
Genus Siphouvigerina Parr, 1950
-
Siphouvigerina hispida (Schwager, 1866)
-
Figure 5: 4a–c
-
1866 Uvigerina hispida Schwager: p. 249, pl. 7, fig. 95
-
1938 Uvigerina rustica Cushman and Edwards: p. 83, pl. 14, fig. 6
-
1980 Euuvigerina hispida (Schwager); Srinivasan and Sharma, pl. 7, figs. 14, 15
-
2002 Neouvigerina hispida (Schwager); Hayward, p. 301, pl. 1, fig. 14
-
2013 Siphouvigerina hispida (Schwager); Patarroyo and Martínez, p. 41
-
2013 Uvigerina hispida Schwager; Holbourn et al. p. 592, fig. 1
Description
Triserial, elongate, nearly fusiform-shaped test with a basal spine. Nearly circular in cross section. Chambers are inflated to sub-globular, gradually increasing in size towards the aperture and separated by depressed sutures. The surface is densely ornamented by acicular to coarse papillae or spines. The aperture is terminal and rounded, with a short neck, a phialine lip and internal tooth plate.
Dimensions
The species length ranges from 175 to 875 µm in the current study.
Stratigraphic range
The species distribution at 752 is from early Serravallian to late Tortonian. At 1139 the species distribution is from Chattian to early Tortonian. At 1168, this species is very rare in the Chattian but it is consistently present from the Aquitanian to early Zanclean.
Remarks
This species is generally larger at Site 1139 than at the other study sites. This species is cosmopolitan (Hayward, 2002; Holbourn et al., 2013) and known from the early Miocene (Aquitanian) to recent. However, it occurs earlier (latest Oligocene) here in the eastern sector of the Indian Ocean.
-
Genus Uvigerina d'Orbigny, 1826
-
Uvigerina auberiana d'Orbigny, 1839a
-
Figure 5: 5a–d
-
1839 Uvigerina auberiana d'Orbigny: p. 106, pl. 2, figs. 23, 24
-
1884 Uvigerina asperula Czjzek; Brady, p. 578, pl. 75, figs. 6, 8
-
1884 Uvigerina asperula var. auberiana Brady, p. 579, pl. 75, fig. 9
-
1978 Uvigerina auberiana (d'Orbigny): Wright, p. 717, pl. 8, fig. 10
-
1984 Uvigerina auberiana (d'Orbigny): Lamb and Miller, p. 26, pl. 4, figs. 1–6
-
2011 Uvigerina auberiana (d'Orbigny): Kuppusamy, p. 69, pl. 13, fig. 9
-
2013 Uvigerina auberiana (d'Orbigny): Holbourn et al., p. 584, figs. 1–3
Description
Test elongate, triserial, fusiform shape. Circular in cross section. Depressed sutures and inflated chambers that gradually increase in size. Small spines usually cover the whole test. Aperture is terminal and rounded, with tubular neck, phialine lip and internal tooth plate.
Dimensions
The species length ranges from 200 to 525 µm in the current study.
Stratigraphic range
The species is only recorded at 752, from Aquitanian to early Zanclean.
Remarks
Some specimens are very densely covered in short spines, ranging all over the test and extending to cover the neck as well. The specimens in this study are relatively elongate (∼2.5–3 times long as broad) similar to the type material rather than the 2:1 ratio described by Holbourn et al. (2013). Cosmopolitan distribution according to Holbourn et al. (2013).
-
Uvigerina peregrina Cushman, 1923
-
Figure 5: 6a–b
-
1923 Uvigerina peregrina Cushman: p. 166, pl. 42, figs. 7–10
-
1926 Uvigerina gardnerae Cushman and Applin: p. 175, pl. 8, figs. 16, 17
-
1984 Uvigerina peregrina Cushman; Lamb and Miller, p. 36-37, pl. 8, figs. 1–3; pl. 9, figs. 1–5
-
1991 Uvigerina peregrina Cushman; Nomura, p. 55, pl. 1, fig. 10
-
2013 Uvigerina peregrina Cushman; Hayward et al., p. 442, pl. 6, fig. 20
Description
Test elongate, triserial. Chambers inflated, separated by depressed sutures. The test is ornamented by parallel, longitudinal costae. Aperture terminal, with phialine lip and short narrow neck.
Dimensions
The species length ranges from 175 to 700 µm in the current study.
Stratigraphic range
The range at 752 and 1139 is Chattian to Tortonian, while at 1168 it ranges from Chattian to early Zanclean.
Remarks
The surface costae are usually non-continuous and parallel. The lower portion of the test is covered in isolated spines, which are randomly oriented; there are also in specimens shown in Lamb and Miller (1984) (pl. 34–37, 8, 9). This species is cosmopolitan (Lamb and Miller, 1984) with records including the Indian Ocean (Nomura, 1991) and New Zealand (Hayward et al., 2013).
-
Order Lagenida Lankester, 1885
-
Superfamily Nodosarioidea Ehrenberg, 1838
-
Family Vaginulinidae Reuss, 1862
-
Subfamily Lenticulininae Chapman, Parr, and Collins, 1934
-
Genus Lenticulina Lamarck, 1804
-
Lenticulina cultrata (de Montfort, 1808)
-
Figure 5: 7a–b
-
1808 Robulus cultratus de Montfort: p. 214
-
1839 Robulina canariensis d'Orbigny: p. 127, pl. 3, figs. 3, 4
-
1846 Robulina similis d'Orbigny: pl. 4, figs. 14, 15
-
2013 Lenticulina cultrata de Montfort; Hewaidy et al., p. 132, pl. 1, figs. 3a, b
-
2016 Lenticulina cultrata de Montfort; Amakrane et al., p. 103, pl. 6, fig. 27
Description
Test planispiral, involute, with peripheral keel. Lenticular-shaped smooth test, circular in outline, biconvex in cross section. Chambers are slightly inflated. Sutures are curved and flush to the test. The primary aperture is radiate and terminal.
Dimensions
The species length ranges from 150 to 1000 µm in the current study.
Stratigraphic range
The species distribution at 752 is Chattian to late Messinian. The species is only present in the early-to-mid Burdigalian at 1139 and it is not recorded at all at 1168.

Figure 6Scale bar is 100 µm. (1a–b) Amphicoryna scalaris, ODP Hole 752A, (1a) 10H-5, 77–81 cm: side view. (1b) 8H-1, 100–105 cm: apertural view. (2a–e) Nuttallides umbonifera, (2a, e) ODP Hole 752A, 4H-5, 64–69 cm: (2a) umbilical view; (2e) edge view. (2b) ODP Hole 752A, 9H-5, 65–70 cm: spiral view. (2c) ODP Hole 1168A, 13X-1, 40–44 cm: spiral view. (2d) ODP Hole 1168A, 14X-4, 45–49 cm: umbilical view. (3a–c) Gyroidinoides soldanii, ODP Hole 1139A, 3R-4, 32–36 cm: (3a) spiral view; (3b) umbilical view; (3c) edge view. (4a–c) Gyroidinoides sp. 1, ODP Hole 1139A, (4a, c) 19R-2, 60.5–64.5 cm: (4a) spiral view; (4c) edge view. (4b) 17R-3, 91–95 cm: umbilical view. (5a–b) Gyroidinoides sp. 2, (5a) ODP Hole 1168A, 32X-4, 80–84 cm: umbilical view. (5b) ODP Hole 1139A, 15R-4, 41–45 cm: spiral view. (6a–d) Oridorsalis umbonatus, (6a–b) ODP Hole 1168A, 8H-1, 100–104 cm: (6a) umbilical view; (6b) spiral view. (6c) ODP Hole 752A, 5H-1, 65–70 cm: spiral view. (6d) ODP Hole 1168A, 8H-1, 100–104 cm: edge view. (7a–c) Osangularia culter, ODP Hole 1168A, 28X-3, 75–79 cm: (7a) umbilical view; (7b) spiral view; (7c) edge view.
Remarks
This species is well-preserved and abundant at 752. It is known from the Indian Ocean (current study) and was noted from the west Pacific (Hanagata and Nobuhara, 2015) so could be considered to have a limited distribution.
-
Subfamily Marginulininae Wedekind, 1937
-
Genus Amphicoryna Schlumberger, in Milne-Edwards, 1881
-
Amphicoryna scalaris (Batsch, 1791)
-
Figure 6: 1a–b
-
1791 Nautilus (Orthoceras) scalaris Batsch: pp. 1, 4, pl. 2, fig. 4a, b
-
1826 Nodosaria longicauda d'Orbigny: p. 254
-
1860 Marginulina falx Jones and Parker: p. 302
-
1884 Nodosaria scalaris (Batsch); Brady, p. 510, pl. 63, figs. 28–31
-
1960 Amphicoryna scalaris (Batsch); Barker, p. 134, pl. 63, figs. 28–31
-
2013 Amphicoryna scalaris (Batsch); Holbourn et al., p. 42, fig. 1
Description
Test elongate, rectilinear, with pronounced basal spine. Chambers are inflated, gradually increasing in size and separated by straight, depressed sutures. The wall is ornamented by long striations or fine ribs many of which are parallel and continuous in appearance. Aperture is terminal and radiate with ring-like ridges around the neck.
Dimensions
The species length ranges from 200 to 750 µm in this study.
Stratigraphic range
At 752 the species ranges from Chattian to Messinian. At 1139 and 1168 this species is very rare, occurring in the mid-Burdigalian at 1139, extending to the early Zanclean at 1168.
Remarks
This species is very well-preserved and recorded in highest abundance at 752. This species occurs worldwide according to Holbourn et al. (2013) and is commonly found in the Indian Ocean.
-
Order Rotaliida Lankester, 1885
-
Superfamily Asterigerinoidea d'Orbigny, 1839a
-
Family Epistomariidae Hofker, 1954
-
Subfamily Nuttallidinae Saidova, 1981
-
Genus Nuttallides Finlay, 1939
-
Nuttallides umbonifera (Cushman, 1933a)
-
Figure 6: 2a–e
-
1933a Pulvinulinella umbonifera Cushman: p. 90, pl. 9, fig. 9
-
1884 Truncatulina pygmaea Hantken; Brady, p. 666, pl. 95, figs. 9, 10
-
1933a Pulvinulinella umbonifera Cushman: p. 90, pl. 9, fig. 9
-
1960 Eponides bradyi Earland; Barker, pl. 95, figs. 9, 10
-
1987 Nuttallides umbonifera (Cushman); Miller and Katz, p. 136, pl. 5, fig. 5
-
1994 Osangulariella umbonifera (Cushman); Jones, p. 99, pl. 95, figs. 9, 10
-
1995 Nuttallides umboniferus (Cushman); Nomura, p. 276, pl. 3, fig. 2
Description
Lenticular, trochospiral form. Unequally biconvex test with a sub-acute peripheral keel. About 12 crescentic chambers in the final whorl, separated by slightly depressed, arcuate to sinuous sutures on the umbilical side and oblique sutures on the spiral side. Aperture is an interiomarginal slit extending from the umbilical boss towards periphery.
Dimensions
The species length ranges from 100 to 750 µm in the current study.
Stratigraphic range
The range of the species is Chattian to Tortonian at 1139 and 752. At 1168 it ranges from Chattian to early Zanclean.
Remarks
This species is characterised by an imperforate umbilical boss. Nuttallides umbonifera (Cushman) is often employed as an indicator of Antarctic Bottom Water (AABW) in the Pacific Ocean and Indian Ocean (Corliss, 1979). This species is long-ranging and distributed worldwide. Mackensen et al. (1990) believed that the species is associated with carbonate corrosive bottom water masses in the Weddell Sea. Distribution worldwide according to Holbourn et al. (2013) and found throughout the Indian Ocean and Southern Ocean (Mackensen, 1992; Nomura, 1995).
-
Superfamily Chilostomelloidea Brady, 1881
-
Family Gavelinellidae Hofker, 1956
-
Subfamily Gavelinellinae Hofker, 1956
-
Genus Gyroidinoides Brotzen, 1942
-
Gyroidinoides soldanii (d'Orbigny, 1826)
-
Figure 6: 3a–c
-
1826 Gyroidina soldanii d'Orbigny: p. 278
-
1846 Gyroidina soldanii d'Orbigny; d'Orbigny, pl. 8, figs. 10–12
-
1978 Gyroidina soldanii d'Orbigny; Wright, p. 715, pl. 5, figs. 7–9
-
1992 Gyroidinoides soldanii d'Orbigny; Mackensen, p. 668, pl. 5, figs. 9, 10
-
1994 Gyroidinoides soldanii d'Orbigny; Jones, p. 106, pl. 107, figs. 6, 7
Description
Test is trochospiral and planoconvex. Umbilical side is involute and convex, and the spiral side is flat and evolute. Rounded periphery. Chambers are inflated and separated by slightly depressed sutures that are straight on umbilical side but curved on spiral side. The surface is smooth. The aperture is a narrow interiomarginal slit, extending from umbilicus to the periphery. The umbilicus is open.
Dimensions
The species length ranges from 100 to 750 µm in this study.
Stratigraphic range
This species occurs in the Chattian at all three sites, ranging to the early Zanclean at 752 and 1168, and the early Tortonian at 1139.
Remarks
Most of the specimens observed in this study have a planar or slightly convex spiral side, and relatively small chambers in the last whorl on the spiral side. Distribution worldwide according to Holbourn et al. (2013). This species is now considered a junior synonym of Hansenisca soldanii (Hayward et al., 2019b) in the World Register of Marine Species (WoRMS).
-
Gyroidinoides sp. 1
-
Figure 6: 4a–c
Description
Test is trochospiral, plano-convex. The spiral side is flat and evolute, while the umbilical side is convex and involute. Chambers are generally inflated separated by slightly depressed sutures on both umbilical and spiral sides. The aperture is an interiomarginal slit extending from umbilicus to periphery. The surface is smooth.
Dimensions
The species length ranges from 100 to 550 µm in this study.
Stratigraphic range
The species is rare at 752 only occurring in the mid-Serravallian and early Zanclean. At 1139 the species ranges from Chattian to early Tortonian. At 1168 it is very rare in the Chattian, with a more common range from early Burdigalian to early Zanclean.
Remarks
The chambers are more elongate and larger on the spiral side than in Gyroidinoides soldanii and it lacks the open, narrow umbilicus of G. soldanii and G. sp. 2. The distribution of the species may be restricted to the current study (southern Indian Ocean). The stratigraphic range overlaps with that of G. soldanii.
-
Gyroidinoides sp. 2
-
Figure 6: 5a–b
Description
The test is trochospiral and planoconvex. Sutures flush to slightly depressed on spiral side, while slightly depressed on umbilical side. Aperture is slit-like extending from umbilicus to periphery. The aperture has a long crescentic flap covering the umbilicus.
Dimensions
The species length ranges from 100 to 400 µm in the current study.
Stratigraphic range
Species ranges from Chattian to mid-Serravallian at 1139 and Chattian to early Zanclean at 1168. This species is not recorded at 752.
Remarks
This species differs from Gyroidinoides soldanii by having larger chambers on the spiral side, which is also more convex. Gyroidinoides sp. 1 has slightly more inflated chambers on the spiral side and a shorter, more curved apertural slit than this species. This species may be restricted to the current study area (southern Indian Ocean).
-
Family Oridorsalidae Loeblich and Tappan, 1984
-
Genus Oridorsalis Andersen, 1961
-
Oridorsalis umbonatus (Reuss, 1851)
-
Figure 6: 6a–d
-
1851 Rotalina umbonata Reuss: p. 75, pl. 5, fig. 35
-
1884 Pulvinulina umbonata (Reuss); Brady, p. 695, pl. 105, fig. 2
-
1884 Truncatulina tenera Brady: p. 665, pl. 95, fig. 11
-
1932 Eponides umbonata (Reuss); Nuttall, p. 26, pl. 6, figs. 4, 5
-
1983 Oridorsalis umbonatus (Reuss); Tjalsma and Lohmann, p. 18, pl. 6, fig. 8
-
1988 Oridorsalis umbonata (Reuss); Whittaker, p. 137, pl. 19, figs. 1–3
-
2011 Oridorsalis umbonatus (Reuss); Kuppusamy et al., p. 59, pl. 8, figs. 23, 24
Description
Test trochospiral, lenticular, unequally biconvex, and nearly circular in outline. The convexity of spiral side is greater than the umbilical side. Involute in umbilical and evolute in spiral side. Six nearly inflated chambers in the last whorl. Sutures usually slightly depressed, curved on umbilical side and radial on spiral side. The test is smooth. The aperture is an interiomarginal slit, extending from periphery to umbilicus.
Dimensions
The species length ranges from 100 to 750 µm in this study.
Stratigraphic range
The stratigraphic distribution of the species is Chattian to early Zanclean at 752 and 1168, Chattian to early Tortonian at 1139.
Remarks
This species generally has six chambers, some specimens possess seven chambers, in the last whorl. This species is easily identifiable and long-ranging from the Paleocene to Holocene, and is common worldwide Holbourn et al. (2013).
-
Genus Osangularia Brotzen, 1940
-
Osangularia culter (Parker & Jones, 1865)
-
Figure 6: 7a–c
-
1865 Planorbulina farcta var. ungeriana subvar. culter Parker and Jones: pp. 382, 421, pl. 19, fig. 1
-
1866 Anomalina bengalensis Schwager, p. 259, pl. 7, fig. 111
-
1884 Truncatulina culter (Parker and Jones); Brady, p. 668, pl. 96, fig. 3
-
1953 Osangularia culter (Parker and Jones); Phleger et al., p. 42, pl. 9, figs. 11–16
-
1960 Osangularia bengalensis (Schwager); Barker, pl. 96, fig. 3
-
1994 Osangularia bengalensis (Schwager); Jones, p. 100, pl. 96, fig. 3
Description
Test is trochspiral, lenticular and unequally biconvex; the umbilical side is more convex than spiral side. The umbilical side is involute, while the spiral side is evolute. Acute periphery with a serrated keel. The slightly inflated nearly 10 chambers, separated by slightly elevated, curved sutures on the spiral side, and straight with slightly depressed sutures on umbilical side. The test is smooth with some perforation in the chambers on spiral side. Aperture is slit-like interiomarginal, extending from umbilicus to periphery.
Dimensions
The species length ranges from 125 to 1000 µm in the current study.
Stratigraphic range
The range of the species is Chattian to early Zanclean at 752, Chattian to early Tortonian at 1139. At 1168 it ranges from Aquitanian to Langhian, and then mid-Tortonian to early Zanclean.
Remarks
Osangularia culter is one of several species dominating intermediate water assemblages in the Miocene (Woodruff, 1985). Distribution cosmopolitan (Holbourn et al., 2013).
-
Superfamily Discorboidea Ehrenberg, 1838
-
Family Rosalinidae Reiss, 1963
-
Genus Gavelinopsis Hofker, 1951
-
Gavelinopsis lobatulus (Parr, 1950)
-
Figure 7: 1
-
1913 Discorbina praegeri (Heron-Allen and Earland): p. 122, pl. 10, figs. 8–10
-
1950 Discorbis lobatulus Parr: p. 354, pl. 13, figs. 23–25
-
1960 Gavelinopsis lobatulus (Parr); Barker, p. 182, pl. 88, figs. 1a–c
-
2011 Gavelinopsis lobatulus (Parr); Kuppusamy et al., p. 52, pl. 5, figs. 2, 3

Figure 7Scale bar is 100 µm. (1) Gavelinopsis lobatulus, ODP Hole 752A, 3H-5, 62–67 cm: umbilical view. (2a–c) Sphaeroidina bulloides, (2a) ODP Hole 752A, 8H-1, 100–105 cm: side view. (2b) ODP Hole 1139A, 8R-1, 45–49 cm: apertural side view. (2c) ODP Hole 1168A, 26X-2, 73–77 cm: apertural side view. (3a–b) Laticarinina pauperata, (3a) ODP Hole 752A, 7H-2, 73–78 cm: umbilical view. (3b) ODP Hole 1139A, 18R-1, 96–100 cm: umbilical view. (4a–b) Epistominella exigua, ODP Hole 1168A, 36X-6, 62.5–66.5 cm: (4a) spiral view; (4b) umbilical view. (5a–b) Astrononion umbilicatulum, ODP Hole 1139A, (5a) 19R-2, 60.5–64.5 cm: umbilical view. (5b) 17R-3, 91–95 cm: edge view. (6a–b) Melonis barleeanus, ODP Hole 1139A, (6a) 4R-2, 105–109 cm: umbilical? view. (6b) 19R-2, 60.5–64.5 cm: edge view. (7a–b) Pullenia bulloides, (7a) ODP Hole 752A, 9H-5, 65–70 cm: edge view. (7b) ODP Hole 1139A, 16R-2, 55–59 cm: edge view. (8a–b) Pullenia quinqueloba, ODP Hole 1168A, 35X-1, 65–69 cm: (8a) edge view; (8b) umbilical view.
Description
Test trochoid. The periphery is thickened and keeled. Five slightly inflated chambers in the last whorl. Nearly flush sutures on spiral side, with slightly depressed sutures on umbilical side. The aperture is an interiomarginal slit extending from umbilicus to periphery, having a narrow lip.
Dimensions
The species length ranges from 125 to 375 µm in the current study.
Stratigraphic range
The stratigraphic distribution is early Langhian to early Zanclean at 752 and mid-Burdigalian to Mid-Serravallian at 1139. This species is not recorded at 1168.
Remarks
Most of the specimens of this species are characterised by an umbilical plug and sharp periphery. Recorded from the Indian Ocean and Southern ocean including the Kerguelen Plateau (Mackensen, 1992) and eastern Indian Ocean (Nomura, 1991). This species is considered a subjective junior synonymy of Gavelinopsis praegeri (Hayward et al., (2019c) in WoRMS).
-
Family Sphaeroidinidae Cushman, 1927
-
Genus Sphaeroidina d'Orbigny, 1826
-
Sphaeroidina bulloides d'Orbigny, 1828 in Deshayes, 1828
-
Figure 7: 2a–c
-
1828 Sphaeroidina bulloides d'Orbigny in Deshayes: p. 267
-
1846 Sphaeroidina austriaca d'Orbigny: p. 284, pl. 20, figs. 19–21
-
1848 Sexloculina haueri Czjzek: p. 149, pl. 13, figs. 35–38
-
1884 Sphaeroidina bulloides d'Orbigny; Brady, p. 620, pl. 84, figs. 1, 2
-
1949 Sphaeroidina ciperana Cushman and Todd: p. 67, pl. 12, fig. 14
-
1949 Sphaeroidina compacta Cushman and Todd: p. 19, pl. 4, fig. 14
-
1949 Sphaeroidina compressa Cushman and Todd: p. 20, pl. 4, figs. 15, 16
-
1949 Sphaeroidina nitida Cushman and Todd: p. 20, pl. 4, fig. 13
-
1953 Sphaeroidina japonica Asano: p. 19, pl. 2, figs. 43–44
-
1985 Sphaeroidina bulloides d'Orbigny; Papp and Schmid, p. 96, pl. 90, figs. 7–12
-
2013 Sphaeroidina bulloides d'Orbigny; Holbourn et al., p. 520, figs. 1–3
Description
Test is sub-globular, usually three chambers in the final whorl. The test is variably coiled. Chambers are globular to hemispherical, separated by slightly depressed sutures. The wall is calcareous, imperforate and smooth. The aperture is a crescentic opening, bordered by a lip, occurring where the three last chambers meet.
Dimensions
The species length ranges from 125 to 725 µm in the current study.
Stratigraphic range
The stratigraphic distribution is Chattian to Zanclean at 752 and 1168, and Chattian to Serravallian at 1139.
Remarks
The larger specimens in the current study have a thick and opaque wall; smaller specimens are thinner-walled and appear translucent. Worldwide distribution according to Holbourn et al. (2013) including previous reports from the eastern Indian Ocean (Nomura, 1991).
-
Superfamily Discorbinelloidea Sigal in Piveteau, 1952
-
Family Discorbinellidae Sigal in Piveteau, 1952
-
Subfamily Discorbinellinae Sigal in Piveteau, 1952
-
Genus Laticarinina Galloway and Wissler, 1927
-
Laticarinina pauperata (Parker & Jones, 1865)
-
Figure 7: 3a–b
-
1865 Pulvinulina repanda Fichtell and Moll var. menardii d'Orbigny, subvar. pauperata Parker and Jones: p. 395, pl. 16, figs. 50–51
-
1884 Pulvinulina pauperata (Parker and Jones); Brady, p. 696, pl. 104, figs. 3–11
-
1942 Laticarinina bulbrooki Cushman and Todd: p. 19, pl. 4, figs. 8, 9
-
1942 Laticarinina crassicarinata Cushman and Todd: p. 18, pl. 4, figs. 11, 12
-
1949 Laticarinina pauperata (Parker and Jones); Bermúdez, p. 309, pl. 23, figs. 43–45
-
1953 Laticarinina pauperata (Parker and Jones); Phleger et al., p. 49, pl. 11, figs. 5, 6
-
1991 Laticarinina pauperata (Parker and Jones); Nomura, p. 55, pl. 1, fig. 6
-
1992 Laticarinina pauperata (Parker and Jones); Mackensen, p. 668, pl. 2, figs. 1, 2
-
2011 Laticarinina pauperata (Parker and Jones); Kuppusamy et al., p. 57, pl. 7, fig. 8
Description
Test planoconvex, slightly trochoid. The spiral side is flat; the umbilical side is slightly convex. Chambers are inflated but becoming progressively more loosely coiled with modest rate of enlargement. Chambers are cuneiform on the umbilical side and reniform on the spiral side, and they are separated by radial and slightly depressed sutures. Chambers on the ventral side are radial, while in the dorsal side is more separated. Large, transparent peripheral keel around whole test, projecting in front of final chamber. Aperture is on the inner margin of last formed chamber.
Dimensions
The species length ranges from 175 to 1300 µm in the current study.
Stratigraphic range
The range of the species at 752 is Chattian to early Burdigalian and then again late Langhian to early Zanclean. At 1139 the species appears in the Chattian and later from the mid-Aquitanian to mid-Serravallian. At 1168, it is recorded from the mid-Aquitanian to mid-Messinian.
Remarks
Most specimens are easily distinguished by the presence of a very broad, transparent keel on the periphery.
Biogeography
This species has previously been reported from the eastern Indian Ocean (Nomura, 1991) and Kerguelen Plateau (Mackensen, 1992) but can be found worldwide (Holbourn et al., 2013).
-
Family Pseudoparrellidae Voloshinova, 1952
-
Subfamily Pseudoparrellinae Voloshinova, 1952
-
Genus Epistominella Husezima and Maruhasi, 1944
-
Epistominella exigua (Brady, 1884)
-
Figure 7: 4a–b
-
1884 Pulvinulina exigua Brady: p. 696, pl. 103, figs. 13–14
-
1951 Pseudoparrella exigua (Brady); Phleger and Parker, p. 28, pl. 15, fig. 6
-
1994 Pseudoparrella exigua (Brady); Loeblich and Tappan, p. 146, pl. 307, figs. 1–7
-
1951 Pulvinulinella exigua (Brady); Hofker, p. 322, figs. 219–221
-
1960 Epistominella exigua (Brady); Barker, pl. 103, figs. 13, 14
-
1992 Epistominella exigua (Brady); Mackensen, p. 668, pl. 4, figs. 5, 6
-
1994 Alabaminoides exiguus (Brady); Jones, p. 103, pl. 103, figs. 13, 14
-
2011 Epistominella exigua (Brady); Kuppusamy et al., p. 50, pl. 4, figs. 11, 12
-
2013 Epistominella exigua (Brady); Holbourn et al., p. 240, figs. 1–6
Description
Test trochoid, unequally biconvex. Slightly convex spiral side, convex umbilical side. The periphery is acute. Five to six moderately inflated chambers in the last whorl, separated by curved, flush sutures on spiral side and curved, depressed sutures on umbilical side. Aperture is an interiomarginal slit on umbilical side extending to the face of the final and largest chamber.
Dimensions
The species length ranges from 100 to 275 µm in the current study.
Stratigraphic range
The species is rare at 752 ranging from early Langhian to early Zanclean. The range is Chattian to early Tortonian at 1139 and to early Zanclean at 1168.
Remarks
The abundance of this species is employed as a proxy for constraining the flux of pulsed organic matter (Holbourn et al., 2013). This species is found worldwide including local to this study in the Timor Sea (Loeblich and Tappan, 1994), Kerguelen Plateau (Mackensen, 1992) and Weddell Sea, Antarctic (Thomas, 1990).
-
Superfamily Nonionoidea Schultze, 1854
-
Family Nonionidae Schultze, 1854
-
Subfamily Astrononioninae Saidova, 1981
-
Genus Astrononion Cushman & Edwards, 1937
-
Astrononion umbilicatulum Uchio, 1952
-
Figure 7: 5a–b
-
1952 Astrononion umbilicatulum Uchio: p. 36, pl. 32, fig. 1
-
1978 Astrononion umbilicatulum Uchio; Wright, p. 710, pl. 1, fig. 12
-
2011 Astrononion umbilicatulum Uchio; Kuppusamy et al., p. 44, pl. 1, figs. 7–9
Description
Planispiral involute test. Periphery rounded, elliptical in lateral view. Chambers large and increasing gradually in size. Approximately seven to eight chambers in the last whorl, separated by curved, thickened radial sutures. Aperture is a narrow interiomarginal, crescentic slit, extending across the periphery to the umbilicus.
Dimensions
The species length ranges from 100 to 350 µm in the current study.
Stratigraphic range
The distribution of the species at 752 is early Aquitanian to early Zanclean and Chattian to early Zanclean at 1168. The range is Chattian to early Tortonian at 1139.
Remarks
The flaps that partially cover the sutures in the last whorl leave a small cavity beneath, which look like secondary chamberlets. This species was originally described from the lower Pleistocene of Japan (Uchio, 1952) but has not been widely reported from elsewhere. It is known from the Indian Ocean (current study), ODP Site 757 (Singh and Gupta, 2004), north-west Atlantic Ocean (Kuppusamy et al., 2011) and the Mediterranean (Wright, 1978).
-
Subfamily Pulleniinae Schwager, 1877
-
Genus Melonis de Montfort, 1808
-
Melonis barleeanus (Williamson, 1858)
-
Figure 7: 6a–b
-
1858 Nonionina barleeana Williamson: p. 32, pl. 3, figs. 68, 69
-
1930 Nonion barleeanum (Williamson); Cushman, p. 11, pl. 4, fig. 5
-
1953 Nonion barleeanum (Williamson); Phleger et al., p. 30, pl. 6, fig. 4
-
1979 Melonis barleeanum (Williamson); Corliss, p. 10, pl. 5, fig. 78
-
1988 Melonis barleeanum (Williamson); Loeblich and Tappan, pl. 696, figs. 5, 6
-
1992 Melonis barleeanus (Williamson); Mackensen, p. 668, pl. 5, fig. 2
-
1997 Melonis barleeanus Williamson, 1858; Bergamin et al., p. 38, pl. 2, figs. 1–9
Description
Test planispiral, bi-umbilicate, involute, compressed test. The umbilicus is open. The test is elliptical, elongated in lateral view, rounded periphery. Ten to 11 flattened chambers, separated by broad, flush sutures. The test is smooth and perforated. The aperture is an interiomarginal slit extending to the umbilicus, possessing an apertural lip.
Dimensions
The species length ranges from 150 to 600 µm in this study.
Stratigraphic range
The stratigraphic distribution of the species at 752 is split into two phases, early Langhian to mid-Serravallian and then again late Messinian to Zanclean. At 1139 it ranges from Chattian to mid-Tortonian, while at 1168 it ranges from Chattian to Zanclean.
Remarks
Forms identified here have a prominent apertural lip distinguishing them from morphologically similar Melonis affinis separated out by some authors (see details in Holbourn et al., 2013). However, we consider the morphologically similar Melonis affinis synonymous with M. barleeanus following molecular analyses by Schweizer (2006).
Biogeography
The species is found worldwide, but Holbourn et al. (2013) suggest that it is not present at very high latitudes. The highest southern-recorded occurrence is on the Kerguelen Plateau by Mackensen (1992), which is supported by the current study observations.
-
Genus Pullenia Parker & Jones in Carpenter et al., 1862
-
Pullenia bulloides (d'Orbigny, 1846)
-
Figure 7: 7a–b
-
1826 Nonionina bulloides d'Orbigny: p. 293
-
1884 Pullenia sphaeroides (d'Orbigny); Brady, p. 615, pl. 84, figs. 12, 13
-
1945 Pullenia bulloides (d'Orbigny); Cushman and Todd, p. 64, pl. 11, fig. 5
-
1949 Pullenia bulloides (d'Orbigny); Bermúdez, p. 276, pl. 21, figs. 28–29
-
1978 Pullenia bulloides (d'Orbigny); Wright, p. 716, pl. 7, fig. 7
-
2011 Pullenia bulloides (d'Orbigny); Kuppusamy et al., p. 62, pl. 9, figs. 18, 19
Description
Test planispiral, involute, globular, sub-circular in outline. Four to five inflated chambers in the last whorl, separated by flush to slightly depressed sutures. Very compact test. Aperture is a narrow interiomarginal, crescentic slit, extending from the periphery to umbilicus.
Dimensions
The species length ranges from 125 to 425 µm in this study.
Stratigraphic range
The stratigraphic distribution of the species at 752 and 1168 is Chattian to Zanclean. At 1139 the species ranges from Chattian to mid-Tortonian.
Remarks
This species usually has a broad periphery and a smooth surface. Up to five chambers are present in the final whorl compared to four or five in the holotype and topotype specimens. Distribution is worldwide (Holbourn et al., 2013) including previous reports from the Indian Ocean by Nomura (1991) and (Singh and Gupta, 2004).
-
Pullenia quinqueloba (Reuss, 1851)
-
Figure 7: 8a–b
-
1851 Nonionina quinqueloba Reuss: p. 71, pl. 5, fig. 31a, b
-
1960 Pullenia subcarinata (d'Orbigny); Barker, pl. 84, figs. 14, 15
-
1949 Pullenia quinqueloba (Reuss); Bermúdez, p. 276, pl. 21, figs. 32, 33
-
1983 Pullenia quinqueloba (Reuss); Tjalsma and Lohmann, p. 36, pl. 16, fig. 2
-
1987 Pullenia quinqueloba (Reuss); Miller and Katz, p. 138, pl. 4, figs. 2a–b
Description
Test planispiral, involute. Sub-globular, slightly compressed test, rounded periphery. Chambers are triangular and inflated. Five chambers present in the final whorl, separated by depressed and radial sutures. Aperture is a narrow, crescentic, interiomarginal slit, extending from periphery to umbilicus.
Dimensions
The species length ranges from 125 to 500 µm in the current study.
Stratigraphic range
The range of the species at 752 is Chattian to Zanclean at 1139 Chattian to mid-Serravallian and at 1168 early Aquitanian to Zanclean.
Remarks
This species is distinguished from five-chambered forms of P. bulloides by the more open coiling and less circular outline. This species is recorded worldwide (Holbourn et al., 2013) but is common in the continental shelf sediments of the Yellow Sea and the East China Sea (Lei and Li, 2016) and in the Indian Ocean (Nomura, 1991; Singh and Gupta, 2004).
-
Superfamily Planorbulinoidea Schwager, 1877
-
Family Cibicididae Cushman, 1927
-
Subfamily Cibicidinae Cushman, 1927
-
Genus Cibicidoides Saidova, 1975
-
Cibicidoides bradyi (Trauth, 1918)
-
Figure 8: 1a–b
-
1918 Truncatulina bradyi Trauth, p. 235
-
1884 Truncatulina dutemplei (d'Orbigny); Brady, p. 665, pl. 95, fig. 5
-
1976 Cibicidoides bradyi (Trauth); Pflum and Frerichs, pl. 3, figs. 6, 7
-
1994 Gyroidina bradyi (Trauth); Jones, p. 99, pl. 95, fig. 5
-
2013 Cibicidoides bradyi Trauth); Holbourn et al., p. 162, figs. 1–6

Figure 8Scale bar is 100 µm. (1a–b) Cibicidoides bradyi, ODP Hole 1139A, (1a) 9R-4, 105–110 cm: umbilical view. (1b) 8R-1, 45–49 cm: spiral view. (2a–c) Cibicidoides mundulus, ODP Hole 752A, 3H-1, 50–55 cm: (2a) umbilical view; (2b) edge view; (2c) spiral view. (3a–c) Cibicidoides wuellerstorfi, (normal) ODP Hole 752A, 7H-2, 73–78 cm: (3a) umbilical view; (3b) edge view; (3c) spiral view. (4a–c) Cibicidoides wuellerstorfi, (smooth) ODP Hole 752A, 5H-1, 65–70 cm: (4a) umbilical view; (4b) edge view; (4c) spiral view. (5a–c) Planulina costata, ODP Hole 752A, 7H-2, 73–78 cm: (5a) spiral view; (5b) edge view; (5c) umbilical view. (6a–b) Eggerella bradyi, ODP Hole 1168A; (6a) 42X-3, 70–74 cm: side view. (6b) 10H-2, 80–84 cm: apertural view.
Description
Test trochospiral, unequally biconvex. The umbilical side is involute, and the spiral side is evolute, more convex. Nine inflated chambers in the last whorl, separated by radial curved sutures in umbilical side, and oblique sutures in the spiral side. The aperture is a slit-like opening with lip extending from periphery to spiral side.
Dimensions
The species length ranges from 100 to 450 µm in this study.
Stratigraphic range
The species first appears in the Chattian and ranges to the Zanclean at 752 and 1168; at 1139 it ranges from Chattian to early Tortonian.
Remarks
Distinguished from Cibicidoides robertsonianus (Brady, 1881), with which it can be confused (Holbourn et al., 2013), by having fewer chambers (typically <10) in the last whorl, smaller test and a more rounded periphery. Recorded worldwide (Holbourn et al., 2013), including from the central Indian Ocean by Boersma (1990). Whilst this species is perhaps most commonly referred to as C. bradyi in the literature (e.g. Holbourn et al., 2013), accepted alternate names include Parrelloides bradyi and Heterolepa bradyi (Hayward et al., 2019d, in WoRMS database).
-
Cibicidoides mundulus (Brady, Parker & Jones, 1888)
-
Figure 8: 2a–c
-
1888 Truncatulina mundula Brady, Parker and Jones: p. 228, pl. 45, fig. 25a–c
-
1953 Cibicides kullenbergi Parker in Phleger et al.: p. 49, pl. 11, figs. 7, 8
-
1955 Cibicidoides mundulus (Brady, Parker and Jones); Loeblich and Tappan, p. 25, fig. 4a–c
-
1960 Cibicidoides mundulus (Brady, Parker and Jones); Barker, pl. 95, fig. 6
-
1992 Cibicidoides mundulus (Brady, Parker and Jones); Mackensen, p. 668, pl. 3, figs. 5–7
Description
Unequally biconvex test, trochospiral. The umbilical side is more convex than the spiral side. Chambers are low, inflated, 10 to 12 chambers in the last whorl, separated by curved sutures. The primary aperture is a long, narrow interiomarginal slit, extending from periphery to spiral side. The test is smooth on the umbilical side and highly perforated on the spiral side.
Dimensions
The species length ranges from 125 to 800 µm in this study.
Stratigraphic range
The stratigraphic distribution of the species at 752 and 1168 is Chattian to Zanclean. At 1139A it ranges from the Chattian to early Tortonian.
Remarks
The specimens here all show a pronounced umbilical boss, as also illustrated by Holbourn et al. (2013, p. 196, pl. 1–8) and (Setoyama and Kaminski, 2015, p. 14, fig. 5, pl. 4a–c). Common species found worldwide (Holbourn et al., 2013), including in the eastern Indian Ocean (Nomura, 1991) and Kerguelen Plateau (Mackensen, 1992).
-
Cibicidoides wuellerstorfi (Schwager, 1866)
-
Figure 8: 3a–c
-
1866 Anomalina wuellerstorfi Schwager: p. 258, pl. 7, figs. 105, 107
-
1884 Truncatulina wuellerstorfi (Schwager); Brady, pp. 627, 662, pl. 93, figs. 8, 9
-
1960 Planulina wuellerstorfi (Schwager); Barker, pl. 93, fig. 9
-
1976 Cibicides wuellerstorfi (Schwager); Pflum and Frerichs, p. 116, pl. 4, figs. 2–4
-
1994 Cibicidoides wuellerstorfi (Schwager); Jones, p. 98, pl. 93, figs. 8, 9
-
2015 Cibicidoides wuellerstorfi (Schwager); Setoyama and Kaminski, p. 14, fig. 5, pl. 10a–c
Description
Planispiral to low trochospiral and plano-convex test. Up to 12 elongated and curved chambers in the last whorl; increasing rapidly in size. Peripheral keel so acute periphery. The sutures are depressed and strongly curved. The wall is coarsely perforated on the spiral side. The aperture is a narrow slit, having a narrow lip extending from the periphery to the spiral side.
Dimensions
The species length ranges from 125 to 1000 µm in the current study.
Stratigraphic range
The stratigraphic distribution of the coarse type at 752 is early Aquitanian to Tortonian and mid-Aquitanian to Zanclean at 1168. At 1139 it ranges from mid-Burdigalian to mid-Serravallian. The smooth type ranges from early Tortonian to Zanclean at 752. At 1139 it ranges from Burdigalian to Serravallian, while at 1168 it ranges from Aquitanian to Zanclean.
Remarks
This species has two different styles of test ornamentation as noted by Nomura (1991) in the eastern Indian Ocean (ODP Hole 754A) – a “smooth” and a “coarse” type. The smooth type (pl. 6, fig. 4a–c) has a smooth umbilical side and is relatively inflated in side view compared to the coarse type, which is highly adorned with nodes on the umbilical side and is more compressed in side view. The smooth type is recorded at all three localities in the current study. The coarse type Cibicidoides wuellerstorfi has previously been assigned to three different genera; Fontobia (Schwager), Planulina Revets (1996) and Cibicidoides. Following the molecular phylogenetic studies of Schweizer (2006) and Schweizer et al. (2009, 2011) the latter genus has now been accepted and applied in the literature (Setoyama and Kaminski, 2015). Worldwide distribution (Holbourn et al., 2013), widely reported in the Indian Ocean (e.g. Boersma, 1990; Nomura, 1991).
-
Family Planulinidae Bermúdez, 1952
-
Genus Planulina d'Orbigny, 1826
-
Planulina costata (Hantken, 1875)
-
Figure 8: 5a–c
-
1875 Truncatulina costata Hantken: p. 73, pl. 9, fig. 2a–c
-
1932 Planulina wuellerstorfi Schwager; Nuttall, p. 31, pl. 4, figs. 14, 15
-
1979 Planulina wuellerstorfi Schwager; Sztràkos, p. 86, pl. 26, fig. 4
-
1934 Planulina marialana Hadley: p. 27, pl. 4, figs. 4–7
-
1941 Planulina palmerae van Bellen: in van Bellen et al., 1941, p. 1144, figs. 7–9
-
1986 Planulina costata (Hantken); van Morkhoven et al., p. 212, pl. 72
-
1991 Planulina costata (Hantken); Nomura, p. 55, pl. 4, fig. 5a–c
Description
Low trochoid test. Spiral side is evolute and flattened, while the umbilical side is involute and slightly convex. Chambers number 10 to 12, with a low inflated shape, separated by slightly to highly curved sutures. The test is smooth on the umbilical side and highly perforated with small pores on the spiral side. Aperture is slit-like, peripheral, with a small lip extending towards the umbilicus.
Dimensions
The species length ranges from 150 to 1150 µm in the current study.
Stratigraphic range
This species only occurs at 752 where it ranges from early Aquitanian to early Serravallian.
Remarks
Van Morkhoven et al. (1986) considered that this species may have evolved from Planulina cocoaensis and that P. costata and Planulina renzi may have evolved separately through the Oligocene and early Miocene. The current species is similar to the smooth form of C. wuellerstorfi in overall test shape and by having a smooth umbilical surface, but differs by having a smaller apertural opening, more compressed test and more irregular periphery. It is similar to Planulina karsteni (Petters and Sarmiento, 1956), but the spiral side of P. karsteni is slightly convex, while in Planulina costata it is plano-convex. Species is widely distributed (van Morkhoven et al., 1986) and has previously been recorded from the eastern Indian Ocean (Nomura, 1991).
-
Order Textulariida Lankester, 1885
-
Superfamily Eggerelloidea Ehrenberg, 1838
-
Family Eggerellidae Cushman, 1937
-
Subfamily Eggerellinae Cushman, 1937
-
Genus Eggerella Cushman, 1933a
-
Eggerella bradyi (Cushman, 1911)
-
Figure 8: 6a–b
-
1911 Verneuilina bradyi Cushman: p. 54, fig. 87
-
1937 Eggerella bradyi (Cushman); Cushman, p. 52, pl. 5, fig. 19
-
1951 Eggerella bradyi (Cushman); Phleger and Parker, p. 6, pl. 3, figs. 1, 2
-
1960 Eggerella bradyi (Cushman); Barker, pl. 47, figs. 4–7
-
1988 Eggerella bradyi (Cushman); Loeblich and Tappan, p. 170, pl. 189, figs. 1, 2
-
1991 Eggerella bradyi (Cushman); Nomura, p. 55, pl. 1, fig. 12a, b
Description
The test is trochoid in the initial portion, subsequently changing to a triserial arrangement. The test is nearly conical. Chambers are highly inflated, separated by depressed sutures. The wall is agglutinated, with a calcareous cement. The aperture is a semicircular slit, with narrow lip, located at the base of the last biggest chamber.
Dimensions
The species length ranges from 125 to 550 µm in the current study.
Stratigraphic range
The species ranges from the Chattian to Zanclean at 752 and 1168. At 1139 it ranges from Chattian to early Tortonian.
Remarks
This species is considered cosmopolitan (Kender et al., 2008) and has previously been recorded in the Miocene of the east Indian Ocean by Nomura (1991) and on the Kerguelen Plateau (Mackensen, 1992).
Benthic foraminiferal assemblages from ODP Sites 752, 1139 and 1168 were identified from the >63 µm fraction for the late Oligocene through to Pliocene. Assemblages are highly diverse comprising more than 200 species at each individual site with between 41 % and 57 % of taxa at any one site, not recorded at the other two, but these taxa represent very low proportions of the overall assemblages. Although there is some faunal similarity between the three sites, in terms of species occurrence, the faunal composition at each site differs through the studied interval and each site responds differently to climatic events through the Miocene. Hole 752A yielded 268 species, of which 143 do not occur at the other two sites; at Hole 1139A 224 species were recorded, 92 of which are only recorded at this site and in Hole 1168A, 331 species are recorded with 189 taxa not present at the two other sites. However, the most abundant taxa in this study occur at all sites, with just a few exceptions. Notably, there appear to be environmentally-cued distributions. For example, Bolivina huneri only occurs at Hole 752 and 1168 but does not occur at 1139. Bolivina dilatata is recorded at 1139 and 1168 but not at 752. Bolivina cf. reticulata and Bolivina viennensis are recorded at 752 but not at the other two sites. Bulimina gibba is recorded at 1139 but not at either 1168 or 752. A full discussion of the assemblage data and palaeoceanographic implications are deferred to a later contribution. Here we selected the 52 most abundant species, each representing at least 1 % of the total fauna encountered in this study, to describe and illustrate providing an important reference work for foraminiferal studies in the southern Indian Ocean.
The data used to compile this paper are available at the University of Birmingham eData repository and can be accessed at https://doi.org/10.25500/edata.bham.00000411 (Ridha et al., 2019).
DR has undertaken all sample processing, data collection and analyses under the supervision of IB and KME. All authors have contributed to the writing of the paper.
The authors declare that they have no conflict of interest.
This research used samples and data provided by the International Ocean Discovery Program (IODP). The authors wish to thank Chris Smart (University of Plymouth) for his useful discussions during this project. We would also like to thank Flavia Fiorini (Khalifa University) and a second, anonymous reviewer for their thoughtful comments on the original submission.
Dana Ridha has been supported by the HCED (The Higher Committee for Education Development in Iraq).
This paper was edited by Laia Alegret and reviewed by Flavia Fiorini and one anonymous referee.
Alve, E. and Murray, J. W.: Temporal variability in vertical distributions of live (stained) intertidal foraminifera, southern England, J. Foramin. Res., 31, 12–24, 2001.
Amakrane, J., Azdimousa, A., Rezqi, H., EL Hammouti, K., EL Ouahabi, M., and Fagel, N.: Paleoenvironment and sequence stratigraphy of the late Miocene from the Guercif basin (Northeastern of Morocco), Bulletin de l'Institut Scientifique, 38, 95–110, 2016.
Andersen, H. V.: Foraminifera of the mudlumps, lower Mississippi River Delta, in: Genesis and Paleontology of the Mississippi River Mudlumps, Louisiana Geol. Survey, Geol. Bull, 35, 1–208, 1961.
Asano, K. J.: Miocene foraminifera from the Noto Peninsula, Ishikawa Prefecture, Tohoku University, Institute of Geological Paleontology Short Papers, 5, 1–21, 1953.
Baldi, K. and Hohenegger, J.: Paleoecology of benthic foraminifera of the Baden-Sooss section (Badenian, Middle Miocene, Vienna Basin, Austria), Geol. Carpathica, 59, 411–424, 2008.
Barker, W. R.: Taxonomic Notes on the species figured by H.B. Brady in his report on the foraminifera dredged by HMS Challenger during the years 1873–1876, Society of Economic Paleontologists Mineralogists, Special Publication, 9, 1–238, 1960.
Batsch, A. I. G. C.: Sechs Kupfertafeln mit Conchylien des Seesandes, gezeichnet und gestochen von AJGK Batsch, University Press, Jena, 6 plates, 1791.
Bergamin, L., Carboni, G., and Bella, L.: Melonis pompilioides (Fichtel & Moll) and Melonis barleeanus (Williamson) from Pliocene, Pleistocene and Holocene sediments of Central Italy, Geologica Romana, Roma, 29–45 pp., 1997.
Bermúdez, P. J.: Tertiary smaller foraminifera of the Dominican Republic, Cushman Laboratory for Foraminiferal Research, 25, 1–322, 1949.
Bermúdez, P. J. J. B. D. G., Venezuela: Estudio sistematico de los Foraminiferos Rotaliformes, Boletin de Geologia, Venezuela, 2, 1–230, 1952.
Boersma, A.: Biostratigraphy and biogeography of Tertiary bathyal benthic foraminifers: Tasman Sea, Coral Sea, and on the Chatham Rise (Deep Sea Drilling Project, Leg 90), Initial reports of the deep sea drilling project, 90, 961–1035, 1986.
Boersma, A.: Late Oligocene to late Pliocene benthic foraminifers from depth traverses in the central Indian Ocean, edited by: Duncan, R. A., Backman, J., Peterson, L. C., et al., College Station, TX (Ocean Drilling Program), 315–379, 1990.
Boltovskoy, E.: Late Cenozoic benthonic foraminifera of the Ninetyeast Ridge (Indian Ocean), Elsevier Oceanography Series, 21, 139–175, 1978.
Boltovskoy, E. and de Kahn, G. G.: Cinco nuevos taxones en Orden Foraminiferida, Comunicaciones des Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” e Instituto Nacional de investigación de las Ciencias Naturales, Hydrobiologia, 2, 43–51, 1981.
Boomgaart, L.: Smaller Foraminifera from Bodjonegoro (Java): Smit and Dontje, 1–175, 1949.
Brady, H. B.: Notes on some of the reticularian Rhizopoda of the Challenger Expedition, Part III, Q. J. Microsc. Sci., 21, 31–71, 1881.
Brady, H. B.: Report on the scientific results of the voyage of the HMS Challenger during the years 1873–1876, Report on the foraminifera dredged by HMS Challenger during the years 1873–1876, Zoology, 9, 1–814, 1884.
Brady, H. B., Parker, W. K., and Jones, T. R.: XI. On some Foraminifera from the Abrohlos Bank, The Transactions of the Zoological Society of London, 12, 211–239, 1888.
Brotzen, F.: Flintrännans och trindelrännans geologi (Öresund): Zusammenfassung, Die Geologie der Flint-und Trindelrinne (Öresund), Norstedt, 34, 1–33, 1940.
Brotzen, F.: Die Foraminiferengattung Gavelinella nov. gen. und die Systematik der Rotaliiformes. Årsbok Sveriges Geologiska Undersokning, Sweden, 36, 1–60, 1942.
Carpenter, W. B., Parker, W. K., and Jones, T. R.: Introduction to the study of the Foraminifera, Ray society, London, 436 pp., 1862.
Chapman, F., Parr, W. J., and Collins, A. C.: Tertiary foraminifera of Victoria, Australia-The Balcombian deposits of Port Phillip, Part lll, Journal of the Linnean Society of London, Zoology, 38, 553–577, 1934.
Corliss, B. H.: Taxonomy of Recent deep- sea benthonic foraminifera from the Southeast Indian Ocean, Micropaleontology, 25, 1–19, 1979.
Cushman, J. A.: A monograph of the foraminifera of the North Pacific Ocean. Part 2. Textulariidae, Bulletin of the United States National Museum, 71, 1–108, 1911.
Cushman, J. A.: The foraminifera of the Atlantic Ocean. Part 3. Textulariidae, Bulletin of the United States National Museum, 104, 1–149, 1922.
Cushman, J. A.: The foraminifera of the Atlantic Ocean. Part 4. Lagenidae, Bulletin of the United States National Museum, 104, 1–228, 1923.
Cushman, J. A.: Some new foraminifera from the Velasco Shale of Mexico, Contributions from the Cushman Laboratory for Foraminiferal Research, 1, 18–23, 1925.
Cushman, J. A.: An outline of a reclassification of foraminifera, Contributions from the Cushman laboratory for foraminiferal research, 3, 1–05, 1927.
Cushman, J. A.: The foraminifera of the Atlantic Ocean, Part 7. Nonionidae, Camerinidae, Peneroplidae and Alveolinellidae, Bulletin of the United States National Museum, 104, 1–79, 1930.
Cushman, J. A.: Some new foraminiferal genera, Contributions from the Cushman Laboratory for Foraminiferal Research, 9, 32–38, 1933a.
Cushman, J. A.: Some new Recent foraminifera from the tropical Pacific, Contributions from the Cushman Laboratory for Foraminiferal Research, 9, 77–95, 1933b.
Cushman, J. A.: Smaller foraminifera from Vitilevu, Fiji, Bulletin of Bernice P. Bishop Museum, 119, 102–142, 1934.
Cushman, J. A.: A monograph of the foraminiferal family Valvulinidae. Special Publications, Cushman Laboratory for Foraminiferal Research, 8, 1–210, 1937.
Cushman, J. A.: Some new foraminifera from the Tertiary of the Island of St. Croix, Contributions from the Cushman Laboratory for Foraminiferal Research, 19, 90–93, 1943.
Cushman, J. A. and Applin, E. R.: Texas Jackson foraminifera, AAPG Bulletin, 10, 154–189, 1926.
Cushman, J. A. and Bermúdez, P. J.: New genera and species of foraminifera from the Eocene of Cuba, Contributions from the Cushman Laboratory for Foraminiferal Research, 12, 27–38, 1936.
Cushman, J. A. and Edwards, P. G.: Astrononion a new genus of the foraminifera, and its species, Contributions from the Cushman laboratory for foraminiferal research, 13, 29–36, 1937.
Cushman, J. A. and Edwards, P. G.: Notes on the Oligocene species of Uvigerina and Angulogerina, Contributions from the Cushman Laboratory for Foraminiferal Research, 14, 74–89, 1938.
Cushman, J. A. and Jarvis, P. W.: Some interesting new uniserial foraminifera from Trinidad, Contributions from the Cushman Laboratory for Foraminiferal Research, 10, 71–75, 1934.
Cushman, J. A. and Moyer, D. A.: Some recent foraminifera from off San Pedro, California, Contributions from the Cushman Laboratory for Foraminiferal Research, 6, 49–62, 1930.
Cushman, J. A. and Parker, F. L.: Bulimina and related foraminiferal genera, Professional Paper of the United States Geological Survey, 210-D, 55–176, 1947.
Cushman, J. A. and Todd, R.: The Recent and fossil species of Laticarinina, Contributions from the Cushman Laboratory for Foraminiferal Research, 18, 14–20, 1942.
Cushman, J. A. and Todd, R.: Miocene foraminifera from Buff Bay, Jamaica, Cushman Laboratory for Foraminiferal Research, Special Publication, 15, 1–73, 1945.
Cushman, J. A. and Todd, R.: The genus Sphaeroidina and its species, Contributions from the Cushman Laboratory for Foraminiferal Research, 25, 11–21, 1949.
Czjzek, J.: Beitrag zur Kenntniss der fossilen Foraminiferen des Wiener Beckens, Naturwissenschaftliche Abhadlungen, Wien, 2, 137–150, 1848.
Debenay, J. P.: A guide to 1,000 foraminifera from Southwestern Pacific: New Caledonia, IRD Editions, Paris, 386 pp., 2012.
De Man, E., Van Simaeys, S., De Meuter, F., King, C., and Steurbaut, E.: Oligocene benthic foraminiferal zonation for the southern North Sea Basin, Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 74, 177–195, 2004.
de Montfort, P. D.: Conchyliologie systématique et classification méthodique des coquilles, F. Schoell, Paris, 1808.
Deshayes, G. P.: Les mollusques [in] Cuvier, Le Règne Animal distribué d'après son organisation, pour servir de base à l'histoire naturelle des animaux et d'introduction à l'anatomie comparée, ed. 2, available at: https://archive.org/stream/lesmollusquesave00desh#page/34/ (last access: 2 July 2019), 310 pp., 1828.
d'Orbigny, A. D.: Tableau méthodique de la classe des Céphalopodes, Annales des Sciences Naturelles 7, 245–314, 1826.
d'Orbigny, A. D.: Foraminifères, in: Histoire physique, politique et naturelle de l'île de Cuba, edited by: Sagra, R. D. L., A. Bertrand, Paris, 1–224, 1839a.
d'Orbigny, A. D.: Voyage dans l'Amérique méridionale: foraminifères, P. Bertrand, Paris and Strasbourg, 86 pp., 1839c.
d'Orbigny, A. D.: Foraminiferes fossiles du Bassin Tertiaire de Vienne (Autriche) 2, Paris: Gide et Camp, 312 pp., 1846.
d'Orbigny, A. D.: Prodrome de paléontologie stratigraphique universelle des animaux mollusques & rayonnés, faisant suite au Cours élémentaire de paléontologie et de géologie stratigraphiques, V. Masson, Paris, 394 pp., 1849–1852.
Eade, J. V.: New Zealand Recent foraminifera of the families Islandiellidae and Cassidulinidae, New Zealand Journal of Marine Freshwater Research, 1, 421–454, 1967.
Earland, A.: Foraminifera. Part III. The Falklands sector of the Antarctic (excluding South Georgia), Discovery Reports, University Press, Cambridge, 1–208, 1934.
Ehrenberg, C. G.: Über dem blossen Auge unsichtbare Kalkthierchen und Kieselthierchen als Hauptbestandtheile der Kreidegebirge, Bericht über die zu Bekanntmachung geeigneten Verhandlungen der Königlichen Preussischen Akademie der Wissenschaften zu Berlin, 192–200, 1838.
Fichtel, L. and Moll, J. P. C.: Testacea Microscopica aliaque Minuta ex Generibus Argonauta et Nautilus ad Naturam Delineat et Descripta, Anton Pichler, Camesina, Vienna, 123 pp., 1798.
Finlay, H. J.: New Zealand foraminifera: key species in stratigraphy – no. 1, T. Roy. Soc. NZ, 68, 504–543, 1939.
Finlay, H. J.: New Zealand foraminifera: key species in stratigraphy – no. 4, T. Roy. Soc. NZ, 69, 448–472, 1940.
Finlay, H. J.: New Zealand Foraminifera: Key Species in Stratigraphy – no. 5, New Zealand Journal of Science And Technology, 28, 259–292, 1947.
Flower, B. and Kennett, J.: Middle Miocene ocean-climate transition: High-resolution oxygen and carbon isotopic records from Deep Sea Drilling Project Site 588A, southwest Pacific, Paleoceanography and Paleoclimatology, 8, 811–843, 1993.
Flower, B. P. and Kennett, J. P.: The middle Miocene climatic transition: East Antarctic ice sheet development, deep ocean circulation and global carbon cycling, Palaeogeogr. Palaeocl., 108, 537–555, https://doi.org/10.1016/0031-0182(94)90251-8, 1994.
Flower, B. P. and Kennett, J. P.: Middle Miocene deepwater paleoceanography in the southwest Pacific: relations with East Antarctic Ice Sheet development, Paleoceanography, 10, 1095–1112, 1995.
Fornasini, C.: Contributo a la conoscenza de le Bulimine adriatiche, Memorie della Reale Accademia delle Scienze dell'Istituto di Bologna, 9, 371–382, 1902.
Fursenko, A.: Osnovnye etapy razvitiya faun foraminifer v geologicheskom proshlom, Trudy Instituta Geologicheskikh Nauk, Akademiia Nauk Belorusskoi SSR, 1, 10–29, 1958.
Galloway, J. J.: A manual of Foraminifera, Principia press, Bloomington, 483 pp., 1933.
Galloway, J. J. and Wissler, S. G.: Correction of names of foraminifera, J. Paleontol., 1, 193–193, 1927.
Glaessner, M. F.: Die Entfaltung der Foraminiferenfamilie Buliminidae, Problemy Paleontologii, Paleontologicheskaya Laboratoriya Moskovskogo Gosudarstvennogo Universiteta, 2, 411–413, 1937.
Griffith, J. W. and Henfrey, A.: The micrographic dictionary: a guide to the examination and investigation of the structure and nature of microscopic objects, J. Van Voorst, London, 845 pp., 1875.
Guérin-Méneville, F. E.: Iconographie du règne animal de G. Cuvier: ou, Représentation d'après nature de l'une des espèces les plus et souvent non encore figurées de chaque genre d'animaux, Paris, London, 48 pp., 1829–1844.
Gümbel, C. W.: Beiträge zur Foraminiferenfauna der nordalpinen, älteren Eocängebilde oder der Kressenberger Nummulitenschichten, Abhandlungen der Mathematisch-Physikalischen Klasse der Königlich Bayerischen Akademie der Wissenschaften, 10, 581–730, 1868.
Guppy, R. J. L.: On some Foraminifera from the Microzoic deposits of Trinidad, West Indies, P. Zool. Soc. Lond., 4, 647–653, 1894.
Gupta, A. K.: Biostratigraphic vs. paleoceanographic importance of Stilostomella lepidula (Schwager) in the Indian Ocean, Micropaleontology, 47–51, 1993.
Gupta, A. K. and Satapathy, S.: Latest Miocene–Pleistocene abyssal benthic foraminifera from west-central Indian Ocean DSDP Site 236: Paleoceanographic and paleoclimatic inferences, Journal of Paleontological Society of India, 45, 33–48, 2000.
Gupta, A. K., Singh, R. K., Joseph, S., and Thomas, E.: Indian Ocean high-productivity event (10–8 Ma): Linked to global cooling or to the initiation of the Indian monsoons?, Geology, 32, 753–756, 2004.
Hadley, W. H.: Some Tertiary foraminifera from the north coast of Cuba, Bulletin of American Paleontology, 20, 1–40, 1934.
Haeckel, E.: Systematische Phylogenie. Entwurf eines Natürlichen Systems der Organismen auf Grund ihrer Stammesgeschichte. Theil l, Systematische Phylogenie der Protisten und Pflanzen, Georg Reimer, Berlin, xv + 400 pp., 1894.
Hanagata, S. and Nobuhara, T.: Illustrated guide to Pliocene foraminifera from Miyakojima, Ryukyu Island Arc, with comments on biostratigraphy, Palaeontologia Electronica, 18, 1–140, 2015.
Hantken, M. V.: Die fauna der Clavulina Szabói-Schichten. Theil I – Foraminiferen, Kaiserlich Ungarische Anstalt, Mitteilungen, Jahrbuch, 4, 1–93, 1875.
Hayward, B. W.: Late Pliocene to middle Pleistocene extinctions of deep-sea benthic foraminifera (“Stilostomella extinction”) in the southwest Pacific, J. Foramin. Res., 32, 274–307, 2002.
Hayward, B. W. and Kawagata, S.: Extinct foraminifera figured in Brady's Challenger Report, J. Micropalaeontol., 26, 171–175, 2005.
Hayward, B. W., Neil, H., Carter, R., Grenfell, H. R., and Hayward, J. J.: Factors influencing the distribution patterns of Recent deep-sea benthic foraminifera, east of New Zealand, Southwest Pacific Ocean, Mar. Micropaleontol., 46, 139–176, https://doi.org/10.1016/S0377-8398(02)00047-6, 2002.
Hayward, B. W., Grenfell, H. R., Sabaa, A., and Hayward, J. J.: Recent benthic foraminifera from offshore Taranaki, New Zealand, New Zealand J. Geolo. Geophys., 46, 489–518, 2003.
Hayward, B. W., Kawagata, S., Grenfell, H. R., Sabaa, A. T., and O'Neill, T.: Last global extinction in the deep sea during the mid-Pleistocene climate transition, Paleoceanography 22, PA3103, https://doi.org/10.1029/2007PA001424, 2007.
Hayward, B. W., Sabaa, A. T., Thomas, E., Kawagata, S., Nomura, R., Schröder-Adams, C., Gupta, A. K., and Johnson, K.: Cenozoic record of elongate, cylindrical, deep-sea benthic foraminifera in the Indian Ocean (ODP Sites 722, 738, 744, 758, and 763), J. Foramin. Res., 40, 113–133, 2010.
Hayward, B. W., Kawagata, S., Sabaa, A., Grenfell, H., Kerckhoven, L. V., Lewandowski, K., and Thomas, E.: The last global extinction (Mid-Pleistocene) of deep sea benthic foraminifera (Chrysalogoniidae, Ellipsoidinidae, Glandulonodosariidae, Plectofrondiculariidae, Pleurostomellidae, Stilostomellidae), their Late Cretaceous-Cenozoic history and taxonomy, Cushman Foundation for Foraminiferal Research, Special Publication, Allen Press, Lawrence, USA, 408 pp., 2012.
Hayward, B. W., Sabaa, A., Grenfell, H., Neil, H., and Bostock, H.: Ecological distribution of Recent deep-water foraminifera around New Zealand, J. Foramin. Res., 43, 415–442, https://doi.org/10.2113/gsjfr.43.4.415, 2013.
Hayward, B. W., Le Coze, F., Vachard, D., and Gross, O.: World Foraminifera Database, Bulimina alazanensis Cushman, 1927, available at: http://www.marinespecies.org/foraminifera/aphia.php?p=taxdetails&id=113032, last access: 7 July 2019a.
Hayward, B. W., Le Coze, F., Vachard, D., and Gross, O.: World Foraminifera Database. Hansenisca soldanii (d'Orbigny, 1826),World Register of Marine Species, available at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=113418, last access: 7 July 2019b.
Hayward, B. W., Le Coze, F., Vachard, D., and Gross, O.: World Foraminifera Database. Gavelinopsis praegeri (Heron-Allen & Earland, 1913), World Register of Marine Species, available at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=113159, last access: 7 July 2019c.
Hayward, B. W., Le Coze, F., Vachard, D., and Gross, O.: World Foraminifera Database. Heterolepa bradyi (Trauth, 1918), World Register of Marine Species, available at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=736954, last access: 7 July 2019d.
Hermelin, J.: The benthic foraminiferal faunas of sites 725, 726, and 728 (Oman Margin, northwestern Arabian Sea), Proceedings of the Ocean Drilling Program. Scientific Results, 117, 55–87, 1991.
Hermoyian, C. S. and Owen, R. M.: Late Miocene‐early Pliocene biogenic bloom: Evidence from low‐productivity regions of the Indian and Atlantic Oceans, Paleoceanography, 16, 95–100, 2001.
Heron-Allen, E. and Earland, A.: Clare Island survey Foraminifera: P. Roy. Irish Acad., 31, 1–188, 1913.
Hewaidy, A. G. A., Sallam, M. M., and Khalifa, M. F.: Miocene calcareous foraminifera of the Nile delta area, Egypt, Egypt. J. Paleontol., 13, 121–171, 2013.
Hilgen, F. J., Lourens, L. J., Van Dam, J. A., Beu, A. G., Boyes, A. F., Cooper, R. A., Krijgsman, W., Ogg, J. G., Piller, W. E., and Wilson, D. S.: Chapter 29 – The Neogene Period, in: The Geologic Time Scale, edited by: Gradstein, F. M., Ogg, J. G., Schmitz, M. D., and Ogg, G. M., Elsevier, Boston, 923–978, 2012.
Hofker, J.: The foraminifera of the Siboga expedition. Part III Siboga-Expeditie, Monographie, IVa, 1–513, 1951.
Hofker, J.: Über die Familie Epistomariidae (Foram.), Palaeontographica Abteilung A, A105, 166–206, 1954.
Hofker, J.: Tertiary foraminifera of coastal Ecuador: Part II, Additional notes on the Eocene species, J. Paleontol., 30, 891–958, 1956.
Holbourn, A., Henderson, A., and Macleod, N.: Atlas of Benthic Foraminifera, John Wiley & Sons, Natural History Museum, Chichester, UK, 642 pp., 2013.
Howe, H. V.: Louisiana Cook Mountain Eocene Foraminifera, Bulletin of the Geological Survey of Louisiana, 14, 1–122, 1939.
Husezima, R. and Maruhasi, M.: A new genus and thirteen new species of foraminifera from the core-sample of Kashiwazaki oil field, Niigata-ken, Journal of the Sigenkagaku Kenkyusho, 1, 391–400, 1944.
Jones, R. W.: The Challenger Foraminifera, Oxford University Press, Oxford, 149 pp., 1994.
Jones, T. R.: The micrographic dictionary; a guide to the examination and investigation of the structure and nature of microscopic objects, edited by: Griffith, J. W. and Henfrey, A., London, 316–320, 1875.
Jones, T. R. and Parker, W. K.: On the rhizopodal fauna of the Mediterranean, compared with that of the Italian and some other Tertiary deposits, Quarterly Journal of the Geological Society, 16, 292–307, 1860.
Kaiho, K.: Eocene to Quaternary benthic foraminifers and paleobathymetry of the Izu-Bonin Arc, Legs 125 and 126, Proceedings of the Ocean Drilling Program, Scientific Results, 126, 285–310, https://doi.org/10.2973/odp.proc.sr.126.137.1992, 1992.
Kaiho, K.: Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean, Geology, 22, 719-722, 1994.
Kaminski, M. A.: The year 2000 classification of the agglutinated Foraminifera, edited by: Bubík, M. and Kaminski, M. A., Proceedings of the Sixth International Workshop on Agglutinated Foraminifera, 237–255, 2004.
Kender, S., Kaminski, M. A., and Jones, R. W.: Early to middle Miocene foraminifera from the deep-sea Congo Fan, offshore Angola, Micropaleontology, 54, 477–568, 2008.
Kuppusamy, M., Gupta, A., and Bhaumik, A.: Distribution of deep-sea benthic foraminifera in the Neogene of Blake Ridge, NW Atlantic Ocean, J. Micropalaeontol., 30, 33–74, 2011.
Lamarck, J. B.: Suite des mémoires sur les fossiles des environs de Paris Annales Muséum National d'Histoire Naturelle, 5, 179–188, 1804a.
Lamb, J. L. and Miller, T. H.: Stratigraphic Significance of Uvigerinid Foraminifers in Western Hemisphere, University of Kansas Paleontological Institute, Paleontological Contributions, 98 pp., 1984.
Lankester, E. R.: Protozoa, in: Encyclopaedia Britannica, 9th edn., Encyclopaedia Britannica, Inc., London, UK, 19, 830–866, 1885.
Lee, J. J.: Phylum Granuloreticulosa (Foraminifera), in: Handbook of Protoctista, edited by: Margulis, L., Corliss, J. O., Melkonian, M., and Chapman, D. J., Jones and Bartlett, Boston, 524–528, 1990.
Lei, Y. and Li, T.: Atlas of Benthic Foraminifera from China Seas: The Bohai Sea and the Yellow Sea, Springer, Berlin, 399 pp., 2016.
Loeblich, A. and Tappan, H.: Revision of some Recent Foraminiferal genera, Smithsonian Miscellaneous Collections, 128, 1–37, 1955.
Loeblich, A. R. and Tappan, H.: Suprageneric classification of the Foraminiferida (Protozoa), Micropaleontology, 30, 1–70, 1984.
Loeblich, A. and Tappan, H.: Foraminiferal genera and their classification Van Nostrand Reinhold, New York, 970 pp., 1988.
Loeblich, A. R. and Tappan, H.: Present status of Foraminiferal Classification, in: Studies in Benthic Foraminifera, edited by: Takayanagi, Y. and Saito, T., Tokai University Press, Tokyo, 93–102, 1992.
Loeblich, A. R. and Tappan, H.: Foraminifera of the Sahul Shelf and Timor Sea, Special Publications of the Cushman Laboratory for Foraminiferal Research, 31, 1–661, 1994.
Mackensen, A.: Neogene benthic foraminifers from the southern Indian Ocean (Kerguelen Plateau): biostratigraphy and paleoecology, edited by: Wise Jr., S. W., Schlich, R., et al., College Station, TX (Ocean Drilling Program), 649–673, 1992.
Mackensen, A., Grobe, H., Kuhn, G., and Fütterer, D. K.: Benthic foraminiferal assemblages from the eastern Weddell Sea between 68 and 73∘ S: Distribution, ecology and fossilization potential, Mar. Micropaleontol., 16, 241–283, https://doi.org/10.1016/0377-8398(90)90006-8, 1990.
Mancin, N., Hayward, B. W., Cobianchi, M., and Lupi, C.: Can the morphology of deep-sea benthic foraminifera reveal what caused their extinction during the mid-Pleistocene Climate Transition?, Marine Micropaleontology, Elsevier, 53–70 pp., 2013.
Marks, P.: A revision of the smaller foraminifera from the Miocene of the Vienna Basin, Contributions from the Cushman Foundation for Foraminiferal Research, 2, 33–73, 1951.
Mazumder, A., Henriques, P., and Nigam, R.: Distribution of benthic foraminifera within oxygen minima zone, off central west coast, India, Gondwana Geol. Mag., 6, 5–10, 2003.
Miller, K. G. and Katz, M. E.: Oligocene to Miocene benthic foraminiferal and abyssal circulation changes in the North Atlantic, Micropaleontology, 33, 97–149, 1987.
Milne-Edwards, A.: Compte rendu sommaire d'une exploration zoologique faite dans l'Atlantique à bord du navire de l'etat “le Travailleur”, Compte Rendu Hebdomadaire des Séances de l' Académie des Sciences, 93, 876–882, 1881.
Neugeboren, J. L.: Die foraminiferen aus der Ordnung der Stichostegier von Ober-Lapugy in Siebenbürgen, Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe, 12, 65-108, available at: http://www.biodiversitylibrary.org/item/104339#page/187/ (last access: 5 July 2019), 1856.
Nishimura, A., Konda, I., Matsuoka, K., Nishida, S., and Ohno, T.: Microfossils of the core sample GDP-11-15 from the Amami Plateau, the northern margin of the Philippines Sea, Memoirs of the Faculty of Science, Kyoto University, Series of geology and mineralogy, 43, 111–129, 1977.
Nomura, R.: Cassidulinidae (foraminiferida) from the uppermost Cenozoic of Japan (part I), The science reports of the Tohoku University, Second series, Geology, 53, 1–101, 1983a.
Nomura, R.: Cassidulinidae (Foraminiferida) from the uppermost Cenozoic of Japan (Part 2), Science Reports of the Tohoku University, Sendai, Series 2 (Geology), 54, 1–93, 1983b.
Nomura, R.: Oligocene to Pleistocene benthic foraminifer assemblages at sites 754 and 756 eastern Indian Ocean, Proceedings Oceanic Drilling Program (ODP), Scientific Results, 121, 31–75, https://doi.org/10.2973/odp.proc.sr.121.139.1991, 1991.
Nomura, R.: Paleogene to Neogene deep-sea paleoceanography in the eastern Indian Ocean: benthic foraminifera from ODP Sites 747, 757 and 758, Micropaleontology, 41, 251–290, 1995.
Nuttall, W. L. F.: Lower Oligocene foraminifera from Mexico, J. Paleontol., 6, 3–35, 1932.
Ohkushi, K. I., Thomas, E., and Kawahata, H.: Abyssal benthic foraminifera from the northwestern Pacific (Shatsky Rise) during the last 298 kyr, Mar. Micropaleontol., 38, 119–147, https://doi.org/10.1016/S0377-8398(99)00040-7, 1999.
O'Neill, T. A., Hayward, B. W., Kawagata, S., Sabaa, A. T., and Grenfell, H. R.: Pleistocene extinctions of deep-sea benthic foraminifera: the South Atlantic record, Palaeontology, 50, 1073–1102, 2007.
Palmer, D. K.: Foraminifera of the Upper Oligocene Cojimar Formation of Cuba, Sociedad Cubana de Historia Natural, 14, 19–35, 113–132, 277–304, 1940.
Palmer, D. K. and Bermúdez, P. J.: Late Tertiary foraminifera from the Matanzas Bay region, Cuba, Memorias de la Sociedad Cubana de Historia Natural Felipe Poey, 9, 237–257, 1936.
Papp, A. and Schmid, M. E.: Die fossilen Foraminiferen des tertiaeren Beckens von Wien: Revision der Monographie Alcide d' Orbigny (1846), Abhandlungen der Geologischen Bundesanstalt, 37, 1–311, 1985.
Parker, F. L.: Distribution of the foraminifera in the North- eastern Gulf of Mexico, Bulletin of the Museum of Comparative Zoology, 111, 453–588, 1954.
Parker, W. K. and Jones, T. R.: On some foraminifera from the North Atlantic and Arctic Oceans, including Davis Straits and Baffin's Bay, Philoso. T. R. Soc., 155, 325–441, 1865.
Parr, W. J.: Victorian and South Australian shallow-water foraminifera, Part II, Proceedings of the Royal Society of Victoria, 44, 1–14, 1932.
Parr, W. J.: Foraminifera, BANZ Antarctic Research Expedition 1929–31, Report, ser, B, 5, 232–392, 1950.
Patarroyo, G. and Martínez, J.: Foraminíferos bentónicos recientes en las aguas profundas de la cuenca de Panamá: Ecología y su posible relación con las corrientes de fondo, Boletín de Investigaciones Marinas y Costeras, 42, 33–58, 2013.
Patterson, R. T.: Abditodendrix, a new foraminiferal genus in the family Bolivinitidae, J. Foramin. Res., 15, 138–140, 1985.
Patterson, R. T.: Four new foraminiferal (Protozoa) genera from the Rio Grande Rise, southwest Atlantic Ocean, Transactions of the American Microscopical Society, 106, 139–148, 1987.
Persico, D., Wise Jr., S. W., and Jiang, S.: Oligocene–Holocene calcareous nannofossil biostratigraphy and diagenetic etch patterns on Quaternary placoliths at ODP Site 1139 on Skiff Bank, Northern Kerguelen Plateau, Proc. ODP, Sci. Results, 1–19, 2003.
Peryt, D.: Foraminiferal record of the Middle Miocene climate transition prior to the Badenian salinity crisis in the Polish Carpathian Foredeep Basin (Central Paratethys), Geol. Q., 57, 141–164, https://doi.org/10.7306/gq.1080, 2013.
Petters, V. and Sarmiento, S. R.: Oligocene and Lower Miocene Biostratigraphy of the Carmen-Zambraon Area, Colombia, Micropaleontology, 2, 7–35, 1956.
Pflum, C. E. and Frerichs, W. E.: Gulf of Mexico deep-water foraminifers, Cushman Foundation for Foraminiferal Research, 125 pp., 1976.
Phleger, F. B. and Parker, F. L.: Ecology of foraminifera, northwest Gulf of Mexico. Pt. II. Foraminifera species, Memoirs of the Geological Society of America, 46, 1–64, 1951.
Phleger, F. B., Parker, F. L., and Peirson, J. F.: North Atlantic foraminifera: Swedish Deep-Sea Exped, Repts., Göteborg, 7, 1–122, 1953.
Piveteau, J.: Traité de Paléontologie, Masson et Cie, Paris, 782 pp., 1952.
Poag, C. W.: Benthic Foraminifera of the Gulf of Mexico: Distribution, ecology, paleoecology, Texas A&M University Press, 244 pp., 2015.
Popescu, G. and Crihan, I.-M.: Middle Miocene foraminifera from Romania: order Buliminida, part II, Acta Palaeontol. Rom., 5, 397–412, 2005.
Rasmussen, T. L.: Systematic paleontology and ecology of benthic foraminifera from the Plio-Pleistocene Kallithea Bay section, Rhodes, Greece, Cushman Foundation Special Publication, 39, 53–157, 2005.
Reiss, Z.: Reclassification of perforate foraminifera, Bulletin of the Geological Survey of Israel, 35, 1–111, 1963.
Renz, H. H.: Stratigraphy and fauna of the Agua Salada group, State of Falcon, Venezuela, Geological Society of America, New York, 219 pp., 1948.
Reuss, A. E.: Neue Foraminiferen aus den Schichten des österreichischen Tertiärbeckens, Denkschriften der Kaiserlichen Akademie der Wissenschaften, 1, 365–390, 1850.
Reuss, A. E.: Ueber die fossilen Foraminiferen und Entomostraceen der Septarienthone der Umgegend von Berlin, Zeitschrift der Deutschen Geologischen Gesellschaft, 3, 49–92, 1851.
Reuss, A. E.: Die Foraminiferen der westphälischen Kreideformation, Sitzungsberichte der mathematisch-naturwissenschaflichen Classe der kaiserlichen Akademie der Wissenschaften, 40, 147–238, 1860.
Reuss, A. V.: Entwurf einer systematischen Zusammenstellung der Foraminiferen, Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften zu Wien, mathematish-naturwissenschaftlichen Classe, 44, 355–396, 1862.
Revets, S. A.: The revision of the genus Buliminellita Cushman and Stainforth, 1947, and Eubuliminella gen. nov, J. Foramin. Res., 23, 141–151, 1993.
Revets, S. A.: The generic revision of five families of rotaliine foraminifera. Part I The Bolivinitidae Cushman, 1927, Cushman Foundation for Foraminiferal Research, Special Publication, 34, 1–55, 1996.
Ridha, D., Boomer, I., and Edgar, K. M.: Latest Oligocene to earliest Pliocene deep-sea benthic foraminifera from ODP Sites 752, 1168 and 1139, southern Indian Ocean, University of Birmingham eData repository, https://doi.org/10.25500/edata.bham.00000411, 2019.
Robertson, B. E.: Systematics and paleoecology of the benthic Foraminiferida from the Buff Bay section, Miocene of Jamaica, Micropaleontology, 44, 1–266, 1998.
Saidova, K. M.: Bentosnye Foraminifery Tikhogo Okeana [Benthonic foraminifera of the Pacific Ocean], Institut Okeanologii P. P. Shirshova, Akademiya Nauk SSSR, Moscow, 875 pp., 1975.
Saidova, K. M.: On an up-to-date system of supraspecific taxonomy of Cenozoic benthonic foraminifera, Akademiya Nauk SSSR, 73 pp., 1981 (in Russian).
Schroder-Adams, C.: Middle Eocene to Holocene Benthic Foraminifer Assemblages from the Kerguelen Plateau (Southern Indian Ocean), edited by: Barron, J., Larsen, B., et al., College Station, Texas, 611–630, 1991.
Schubert, R. J.: Die Miocäne Foraminiferen fauna von Karwin (Oestern Schlesien), Lotos, 19, 211–247, 1899.
Schultze, M. S.: Über den Organismus der Polythalamien (Foraminiferen), nebst Bermerkungen über die Rhizopoden im Allgemeinen, Leipzig, 68 pp., 1854.
Schwager, C.: Fossile Foraminiferen von Kar Nikobar, Reise der Österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter den Befehlen des Commodore B. von Wüllerstorf-Urbair, 2, 187–268, 1866.
Schwager, C.: Quadro del proposto sistema di classificazione dei foraminiferi con guscio, Bolletino R. Comitato Geologico d'Italia, 8, 18–27, 1877.
Schweizer, M.: Evolution and molecular phylogeny of Cibicides and Uvigerina (Rotaliida, Foraminifera), Geologica Ultraiectina, 261, 1–167, 2006.
Schweizer, M., Pawlowski, J., Kouwenhoven, T., and van der Zwaan, B.: Molecular phylogeny of common Cibicidids and related Rotaliida (Foraminifera) based on small subunit rDNA sequences, J. Foramin. Res., 39, 300–315, 2009.
Schweizer, M., Fontaine, D., and Pawlowski, J.: Phylogenetic position of two Patagonian Cibicididae (Rotaliida, foraminifera): Cibicidoides dispars (d'Orbigny, 1839) and Cibicidoides variabilis (d'Orbigny, 1826), Revue de Micropaléontologie, 54, 175–182, 2011.
Seguenza, G.: Le formazioni terziarie nella provincia di Reggio (Calabria), Memorie della Classe di Scienze Fisiche Matematiche e Naturali della Regia Accademia del Lincei, 3, 1–445, 1880.
Setoyama, E. and Kaminski, M.: Neogene Benthic Foraminifera from the southern Bering Sea (IODP Expedition 323), Palaeontol. Electron., 18, 1–30, 2015.
Shipboard Scientific Party: Site 752, College Station, TX (Ocean Drilling Program), 359–453, 1989.
Shipboard Scientific Party: Site 1139, College Station, TX (Ocean Drilling Program), 1–213, 2000.
Shipboard Scientific Party: Site 1168, College Station, TX (Ocean Drilling Program), 1–170, 2001.
Silvestri, A.: Revisione di fossili della Venezia e della Venezia Giulia, Atti dell'Accademia Scientifica Veneto–Trentino–Istriana, Padova (ser. 3), 14, 7–12, 1924.
Silvestri, O.: Saggio di studi sulla fauna microscopia fossile appartenente al terreno subappenino italiano. Mem. I – monografia delle Nodosarie, Academia Gioenia Scienze Naturali Catania, 3, 1–108, 1872.
Singh, R. K. and Gupta, A. K.: Late Oligocene–Miocene paleoceanographic evolution of the southeastern Indian Ocean: evidence from deep-sea benthic foraminifera (ODP Site 757), Mar. Micropaleontol., 51, 153–170, https://doi.org/10.1016/j.marmicro.2003.10.003, 2004.
Singh, R. K., Gupta, A. K., and Das, M.: Paleoceanographic significance of deep-sea benthic foraminiferal species diversity at southeastern Indian Ocean Hole 752A during the Neogene, Palaeogeogr. Palaeocl., 361, 94–103, 2012.
Smart, C. W., Thomas, E., and Ramsay, A. T.: Middle–late Miocene benthic foraminifera in a western equatorial Indian Ocean depth transect: paleoceanographic implications, Palaeogeogr. Palaeocl., 247, 402–420, 2007.
Srinivasan, M. and Sharma, V.: Schwager's Car Nicobar Foraminifera in the Reports of the Novara Expedition: a revision, Today and Tomorrow Publisher, New Delhi, 83 pp., 1980.
Stainforth, R.: Classification of uniserial calcareous Foraminifera, Contributions from the Cushman Foundation for Foraminiferal Research, 3, 6–14, 1952.
Stickley, C., Brinkhuis, H., McGonigal, K., Chaproniere, G., Fuller, M., Kelly, D., Nürnberg, D., Pfuhl, H., Schellenberg, S., and Schönfeld, J.: Late Cretaceous–Quaternary biomagnetostratigraphy of ODP Sites 1168, 1170, 1171, and 1172, Tasmanian Gateway, Proceedings of the Ocean Drilling Program, Scientific Results, 1–57, 2004.
Sztràkos, K.: La stratigraphie, paléoécologie, paléogéographie et les foraminifères de l'Oligocène du nord-est de la Hongrie, Éditions du Centre national de la recherche scientifique, Paris, 95 pp., 1979.
Sztràkos, K. J. R. D. M.: Les foraminifères de l'Eocène du Bassin de l'Adour (Aquitaine, France): biostratigraphie et taxinomie, Revue de micropaléontologie, 43, 71–172, 2000.
Thalmann, H. E.: Mitteilungen über Foraminiferen III, Eclogae Geologicae Helvetiae, 30, 337–356, 1937.
Thomas, E.: Late Cretaceous through Neogene deep-sea benthic foraminifera (Maud Rise, Weddell Sea, Antarctica), Proceedings Oceanic Drilling Program (ODP), Scientific Results, 113, 571–594, 1990.
Tjalsma, R.: Eocene to Miocene benthic foraminifera from Deep-Sea Drilling Project Site-516, Rio-Grande Rise, South Atlantic, Initial Reports of the Deep Sea Drilling project, 72, 731–755, 1983.
Tjalsma, R. C. and Lohmann, G. P.: Paleocene-Eocene bathyal and abyssal benthic foraminifera from the Atlantic Ocean Micropaleontology, Special Publication 4, 1–90, 1983.
Todd, R.: Smaller Foraminifera, in: Geology of Saipan, Mariana Islands. Part 3. Palaeontology, Professional Papers U.S. Geological Survey, Washington, 265–320, 1957.
Trauth, F.: Das Eozanvorkommen bei Radstadt im Pongau und seine Beziehungen zu den gleichalterigen Ablagerungen bei Kirchberg am Wechsel und Wimpassing am Leithagebirge, Kaiserlichen Akademie der Wissenschaften in Wein, Mathematisch-Naturwissenschaftliche Classe, 95, 171–278, 1918.
Uchio, T.: Foraminiferal assemblages from Hachijo Island, Tokyo Prefecture, with descriptions of some new genera and species, Japanese Journal of Geology and Geography, 22, 145–159, 1952.
van Bellen, R. C., Rutgers, J. G., Soest, J. V., Witt Puyt, J. F. C. D., and Rutgers, A. C.: Smaller Foraminifera from the Lower Oligocene of Cuba, Koninklijke Nederlandse Akademie van Wetenschappen Proceedings, 9, 1140–1146, 1941.
Van Morkhoven, F. P., Berggren, W. A., Edwards, A. S., and Oertli, H.: Cenozoic cosmopolitan deep-water benthic foraminifera, Elf Aquitaine, Pau, France, 421 pp., 1986.
Vella, P.: Studies in New Zealand Foraminifera; Part I- Foraminifera from Cook Strait. Part II – Upper Miocene to Recent Species of the Genus Notorotalia, New Zealand Geological Survey Paleontological Bulletin, 28, 1–64, 1957.
Voloshinova, N. A.: Pseudoparellinae, p. 80, in: Iskopaemye Foraminifery SSSR. Nonionidy, Kassidulinidy I Khilostomellidy, edited by: Voloshinova, N. A., and Dain, L. G., Trudy Vsesoyuznogo Neftyanogo Nauchnoissledovatel'skogo Geologo-razvedochnogo Instituta (VNIGRI), New Series, 1–151, 1952 (in Russian).
Voloshinova, N. A.: Uspekhi mikropaleontologii v dele izucheniya vnutrennego stroeniya foraminifer, Trudy Pervogo Seminara po Mikrofaune, 48–87, 1960.
Weatherall, P., Marks, K. M., Jakobsson, M., Schmitt, T., Tani, S., Arndt, J. E., Rovere, M., Chayes, D., Ferrini, V., and Wigley, R.: A new digital bathymetric model of the world's oceans, Earth Space Sci., 2, 331–345, https://doi.org/10.1002/2015ea000107, 2015.
Wedekind, R.: Einführung in die Grundlagen der historischen Geologie: Mikrobiostratigraphie. Die Korallen-u. Foraminiferenzeit, Enke, Stuttgart, 136 pp., 1937.
Whittaker, J. E.: Benthic Cenozoic Foraminifera from Ecuador: Taxonomy and Distribution of Smaller Benthic Foraminifera from Coastal Ecuador (Late Oligocene-Late Pliocene), British Museum of Natural, 194 pp., 1988.
Williamson, W. C.: On the recent foraminifera of Great Britain, Ray Society, 107 pp., 1858.
Woodruff, F.: Changes in Miocene deep-sea benthic foraminiferal distribution in the Pacific Ocean: relationship to paleoceanography, Geological Society of America Memoir, 163, 131–175, 1985.
Wright, R.: Neogene benthic foraminifera from DSDP leg 42A, Mediterranean Sea. Init. Rept, DSDP, 42, 709–726, 1978.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K.: Trends, rhythms, and aberrations in global climate 65 Ma to present, Science, 292, 686–693, 2001.